

Parasaurolophus (vyslovováno /ˌpærəsɔˈrɒləfəs/ PARR-.ə-saw-ROL-ə-fəs, běžně také /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; ve významu „blízký chocholatý ještěr“ v odkazu na Saurolophus) je rod ornitopodního dinosaura z období pozdní křídy na území dnešní Severní Ameriky, asi před 76-73 miliony let. Byl to býložravec, který se pohyboval jako dvounožec i čtyřnožec. Rozpoznávají se tři druhy: P. walkeri (typový druh), P. tubicen a krátkonohý P. cyrtocristatus. Pozůstatky jsou známy z Alberty v Kanadě a z Nového Mexika a Utahu v USA. Poprvé byl popsán v roce 1922 Williamem Parksem z lebky a částečné kostry v Albertě.
Parasaurolophus je hadrosaurid, součást rozmanité čeledi křídových dinosaurů známých řadou bizarních ozdob hlavy. Tento rod je známý pro svůj velký, propracovaný lebeční hřeben, který v největším rozsahu tvoří dlouhou zakřivenou trubici vyčnívající z lebky směrem vzhůru a dozadu. Charonosaurus z Číny, který mohl být jeho nejbližším příbuzným, měl podobnou lebku a možná i podobný hřeben. O hřebenu vědci hodně diskutovali; panuje shoda, že mezi hlavní funkce patřilo vizuální rozpoznávání druhu i pohlaví, akustická rezonance a termoregulace. Patří mezi vzácnější kachnozobé, je znám pouze z několika dobrých exemplářů.
Parasaurolophus patří mezi nejznámější dinosaury a je často zobrazován v televizních pořadech, filmech, knihách, zboží a dalších médiích souvisejících s dinosaury. Alice Perry
Popis

Velikost Parasaurolophus walkeri ve srovnání s člověkem.
Stejně jako u většiny dinosaurů je kostra Parasaurolofa neúplně známá. Délka typového exempláře P. walkeri se odhaduje na 9,5 metru. Jeho lebka je včetně hřebene dlouhá asi 1,6 metru, zatímco typová lebka P. tubicen je dlouhá přes 2,0 metru, což svědčí o větším zvířeti. Jeho hmotnost se odhaduje na 2,5 tuny (2,7 tuny). Jeho jediná známá přední končetina byla na hadrosaurida poměrně krátká, s krátkou, ale širokou lopatkou. Stehenní kost měří u P. walkeri 103 cm a ve srovnání s jinými hadrosauridy je na svou délku robustní. Silně stavěná byla také horní část paže a pánevní kosti.
Stejně jako ostatní hadrosauridi byl schopen chůze buď po dvou, nebo po čtyřech nohách. Pravděpodobně dával přednost shánění potravy na čtyřech nohách, ale běhal po dvou. Nervové trny obratlů byly vysoké, jak bylo u lambeosaurinů běžné; nejvyšší nad kyčlemi zvyšovaly výšku hřbetu. U P. walkeri jsou známy otisky kůže, které ukazují rovnoměrné hlízovité šupiny, ale žádné větší struktury.
Parasaurolophus walkeri s detailem skalpování.
Nejnápadnějším znakem byl lebeční hřeben, který vyčníval ze zadní části hlavy a byl tvořen premaxillou a nosními kostmi. Typový exemplář P. walkeri má zářez v neurálních trnech poblíž místa, kde by hřeben narážel na zátylek, ale to může být patologie vlastní tomuto jedinci. William Parks, který rod pojmenoval, předpokládal, že od hřebene k zářezu vede vaz, který podpírá hlavu. Ačkoli se tato myšlenka zdá nepravděpodobná, Parasaurolophus je někdy obnovován s kožním lalokem od hřebene ke krku.
Hřeben byl dutý, s výraznými trubicemi vedoucími z každé nozdry na konec hřebene, než obrátily směr a směřovaly zpět dolů po hřebeni a do lebky. Trubice byly nejjednodušší u P. walkeri a složitější u P. tubicen, kde některé trubice byly slepé a jiné se setkávaly a oddělovaly. Zatímco P. walkeri a P. tubicen měli dlouhý hřeben s jen mírným zakřivením, P. cyrtocristatus měl krátký hřeben s kruhovitějším profilem.
Klasifikace
Jak napovídá jeho jméno, Parasaurolophus byl původně považován za blízce příbuzného Saurolophus, protože měl na povrchu podobný hřeben. Brzy však byl přehodnocen na člena lambeosaurní podčeledi hadrosauridů – Saurolophus je hadrosaurin. Obvykle je interpretován jako samostatná odnož lambeosaurinů, odlišná od korythosaura, hypakrosaura a lambeosaura s helmovým hřebenem. Jeho nejbližším známým příbuzným se zdá být Charonosaurus, lambeosaurin s podobnou lebkou (ale zatím bez kompletního hřebene) z amurské oblasti severovýchodní Číny, a oba mohou tvořit klad Parasaurolophini. P. cyrtocristatus s krátkým, kulatějším hřebenem může být nejzákladnějším ze tří známých druhů Parasaurolophus, nebo může představovat subadultní či samičí exempláře P. tubicen.
Historie
Objevení a pojmenování
Odlitek neúplného typového exempláře Parasaurolophus walkeri ve Varšavě.
Význam jména Parasaurolophus je „blízký chocholatý ještěr“ a je odvozen z řeckého para/παρα „vedle“ nebo „blízko“, saurus/σαυρος „ještěr“ a lophos/λοφος „chochol“. Vychází z nálezu ROM 768, lebky a částečné kostry, které chybí většina ocasu a zadní končetiny pod koleny a které byly nalezeny terénní skupinou z Torontské univerzity v roce 1920 poblíž Sand Creek u řeky Red Deer v kanadské Albertě. Tyto horniny jsou dnes známy jako svrchnokřídová formace Dinosaur Park z období kampánu. William Parks pojmenoval exemplář P. walkeri na počest sira Byrona Edmunda Walkera, předsedy správní rady Královského ontarijského muzea. Pozůstatky rodu Parasaurolophus jsou v Albertě vzácné, z (pravděpodobně) dinosauří formace pochází pouze jedna další částečná lebka a třem exemplářům z Dinosaur Park chybí lebky, které by mohly patřit k tomuto rodu. V některých faunistických seznamech je zmínka o možném materiálu P. walkeri ve formaci Hell Creek v Montaně, horninové jednotce pozdně maastrichtského stáří. Tento výskyt Sullivan a Williamson ve svém přehledu rodu z roku 1999 nezaznamenali a jinde nebyl blíže rozveden.
V roce 1921 získal Charles H. Sternberg částečnou lebku (PMU.R1250) z dnes o něco mladší formace Kirtland v okrese San Juan v Novém Mexiku. Tento exemplář byl zaslán do švédské Uppsaly, kde jej Carl Wiman v roce 1931 popsal jako druhý druh P. tubicen. Specifické epiteton je odvozeno z latinského tǔbǐcěn „trubač“. Druhá, téměř kompletní lebka P. tubicen (NMMNH P-25100) byla nalezena v Novém Mexiku v roce 1995. Na základě počítačové tomografie této lebky zpracovali Robert Sullivan a Thomas Williamson v roce 1999 tento rod monograficky, přičemž se zabývali aspekty jeho anatomie a taxonomie a funkcí jeho hřebene. Williamson později publikoval nezávislý přehled pozůstatků, v němž s taxonomickými závěry nesouhlasil.
John Ostrom v roce 1961 popsal další kvalitní exemplář (FMNH P27393) z Nového Mexika jako P. cyrtocristatus. Obsahuje částečnou lebku s krátkým zaobleným hřebenem a většinu postkraniálního skeletu s výjimkou nohou, krku a částí ocasu. Jeho specifické jméno je odvozeno z latinského curtus „zkrácený“ a cristatus „chocholatý“. Exemplář byl nalezen buď ve svrchní části Fruitlandské formace, nebo, což je pravděpodobnější, na bázi nadložní Kirtlandské formace. Areál tohoto druhu byl rozšířen v roce 1979, kdy David B. Weishampel a James A. Jensen popsali částečnou lebku s podobným hřebenem (BYU 2467) z Kaiparowitsovy formace z období kampánu v Garfield County v Utahu. Od té doby byla v Utahu nalezena další lebka s krátkou/kulatou morfologií hřebene P. cyrtocristatus.
Druh
Odlitek lebky Parasaurolophus walkeri, Natural History Museum, London.
Odlitek lebky Parasaurolophus cyrtocristatus, Field Museum of Natural History.
Typ druhu P. walkeri z Alberty je znám z jediného exempláře. Od druhu P. tubicen se liší jednoduššími trubicemi v hřebeni a od druhu P. cyrtocristatus dlouhým, nezaobleným hřebenem a delším ramenem než předloktím.
P. tubicen, z Nového Mexika, je znám z pozůstatků nejméně tří jedinců. Je to největší druh, má složitější průduchy v hřebeni než P. walkeri a delší, rovnější hřeben než P. cyrtocristatus.
P. cyrtocristatus, z Nového Mexika a Utahu, je znám ze tří možných exemplářů. Je to nejmenší druh s krátkým zaobleným hřebenem. Jeho malá velikost a tvar hřebene vedly několik vědců k domněnce, že představuje mláďata nebo samice druhu P. tubicen, který pochází zhruba ze stejné doby a ze stejného útvaru v Novém Mexiku. Jak poznamenal Thomas Williamson, typový materiál P. cyrtocristatus je asi 72 % velikosti P. tubicen, což se blíží velikosti, při níž se podle výkladu u ostatních lambeosaurinů začíná projevovat definitivní pohlavní dimorfismus v hřebeni (~ 70 % velikosti dospělého jedince). Toto stanovisko bylo v nedávných přehledech lambeosaurinů odmítnuto.
Paleobiology
Paleoecology
Gorgosaurus (vlevo) útočí na Parasaurolophus cyrtocristatus.
Parasaurolophus walkeri z formace Dinosaur Park byl členem rozmanité a dobře zdokumentované fauny prehistorických zvířat, včetně známých dinosaurů, jako jsou rohatý Centrosaurus, Styracosaurus a Chasmosaurus; kolegové kachnozobí Prosaurolophus, Gryposaurus, Corythosaurus a Lambeosaurus; tyranosaurid Gorgosaurus; a obrnění Edmontonia a Euoplocephalus. Byl vzácnou součástí této fauny. Formace dinosauřího parku je interpretována jako nízkoreliéfové prostředí řek a záplavových oblastí, které se postupem času stalo bažinatějším a ovlivněným mořskými podmínkami, jak západní vnitrozemská mořská cesta postupovala na západ. Klima bylo teplejší než v dnešní Albertě, bez mrazů, ale s vlhčími a suššími obdobími. Dominantními rostlinami v korunách stromů byly zřejmě jehličnany, v podrostu kapradiny, stromové kapradiny a nahosemenné rostliny.
Novomexické druhy sdílely své prostředí s velkým sauropodem alamosaurem, kachnozobým kritosaurem, rohatým pentaceratopsem, obrněným nodocefalosaurem, saurornitholestem a v současnosti nepojmenovanými tyranosauridy. Kirtlandská formace je interpretována jako říční náplavy, které se objevily po ústupu Západního vnitřního moře. Dominantními rostlinami byly jehličnany a chasmosaurinní rohatí dinosauři byli zřejmě běžnější než hadrosauridi.
Jídlo
Parasaurolophus byl jako hadrosaurid velký bipední/kvadrupední býložravec, živící se rostlinami s propracovanou lebkou, která umožňovala mlecí pohyby analogické žvýkání. Jeho zuby se neustále vyměňovaly a byly zabaleny do zubních baterií, které obsahovaly stovky zubů, z nichž jen relativní hrstka byla vždy v provozu. K ořezávání rostlinného materiálu používal zobák, který byl v čelistech držen orgánem podobným líci. Krmení probíhalo ze země až do výšky asi 4 metrů. Jak poznamenal Bob Bakker, lambeosaurini mají užší zobák než hadrosaurini, z čehož vyplývá, že Parasaurolophus a jeho příbuzní se mohli živit selektivněji než jejich protějšky s širokým zobákem a bez hřebene.
Lebeční hřeben
O tom, jaké funkce plnil lebeční hřeben Parasaurolofa, bylo vysloveno mnoho hypotéz, většina z nich však byla zdiskreditována. Nyní se předpokládá, že mohl mít několik funkcí: vizuální zobrazení pro identifikaci druhu a pohlaví, zesílení zvuku pro komunikaci a termoregulaci. Není jasné, která z nich byla v jakém období vývoje hřebene a jeho vnitřních nosních průduchů nejvýznamnější.
Rozdíly mezi druhy a růstovými stádii
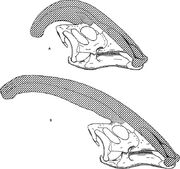
Diagram porovnávající nariální hřebeny Parasaurolophus cyrtocristatus (a) a Parasaurolophus walkeri (b).
Stejně jako u ostatních lambeosaurinů se předpokládá, že lebeční hřeben Parasaurolophus se měnil s věkem a u dospělých jedinců byl pohlavně dimorfním znakem. James Hopson, jeden z prvních badatelů, kteří popsali hřeben lambeosaurinů z hlediska těchto rozdílů, předpokládal, že P. cyrtocristatus s malým hřebenem byl samičí formou P. tubicen. Thomas Williamson se domníval, že jde o juvenilní formu. Ani jedna z těchto hypotéz se nestala obecně přijímanou. Vzhledem k tomu, že je známo pouze šest dobrých lebek a jedna juvenilní mozkovna, další materiál pomůže tyto potenciální vztahy objasnit. Williamson poznamenal, že v každém případě měl juvenilní Parasaurolophus pravděpodobně malé, zaoblené hřebeny jako P. cyrtocristatus, které pravděpodobně rostly rychleji, jak se jedinci blížili dospělosti. Nedávné opětovné studium juvenilního mozkovny dříve přiřazené Lambeosaurovi a nyní přiřazené Parasaurolofovi poskytuje důkaz, že malý trubicovitý hřeben byl u juvenilů přítomen. Na tomto exempláři se zachovalo malé vyklenutí čelních kostí směrem nahoru, které bylo podobné, ale menší než u dospělých jedinců; u dospělých jedinců tvořily čelní kosti platformu, která podpírala základnu hřebene. Tento exemplář také naznačuje, že růst hřebene u parasaurolofa a obličejový profil juvenilních jedinců se lišil od modelu korythosaurus-hypakrosaurus-lambeosaurus, částečně proto, že hřeben parasaurolofa postrádá tenký kostěný „coxcomb“, který tvoří horní část hřebene ostatních tří lambeosaurinů.
Zavržené hypotézy o funkci
Mnoho prvních návrhů se zaměřovalo na adaptace pro vodní způsob života v návaznosti na hypotézu, že hadrosauridi byli obojživelníci, což byl běžný směr uvažování až do 60. let 20. století. Tak Alfred Sherwood Romer navrhoval, že sloužila jako šnorchl, Martin Wilfarth, že byla nástavcem pro pohyblivý chobot, který sloužil jako dýchací trubice nebo ke sběru potravy, Charles M. Sternberg, že sloužila jako vzdušnice, aby se voda nedostala do plic, a Ned Colbert, že sloužila jako zásobárna vzduchu pro delší pobyt pod vodou.
Další návrhy byly spíše fyzikální povahy. Jak bylo zmíněno výše, William Parks navrhoval, že byla spojena s obratli pomocí vazů nebo svalů a pomáhala při pohybu a podpírání hlavy. Othenio Abel navrhl, že se používala jako zbraň v boji mezi příslušníky stejného druhu, a Andrew Milner navrhl, že mohla sloužit jako deflektor listí, podobně jako hřeben přilby (nazývaný „casque“) kasuára. Podle dalších návrhů bylo hlavní funkcí uložení specializovaných orgánů. Halszka Osmólska navrhla, že se v něm nacházely solné žlázy, a John Ostrom předpokládal, že se v něm nacházely rozšířené oblasti pro čichovou tkáň a mnohem lepší čich lambeosaurinů, kteří neměli žádné zjevné obranné schopnosti. Jedním z neobvyklých návrhů, který vyslovil kreacionista Duane Gish, je, že v hřebeni se nacházely chemické žlázy, které mu umožňovaly vrhat na nepřátele proudy chemického „ohně“, podobně jako to dělá dnešní brouk bombardér.
Většina těchto hypotéz byla zdiskreditována nebo zamítnuta. Například na konci hřebene není žádný otvor pro funkci šnorchlu. Nejsou zde žádné svalové jizvy pro chobot a je pochybné, že by ho živočich se zobákem potřeboval. Jako navrhovaný vzdušný uzávěr by nezadržoval vodu. Navrhovaná vzduchová nádrž by byla pro zvíře velikosti parasaurolofa nedostatečná. Ostatní hadrosauridi měli velké hlavy, aniž by potřebovali velké duté hřebeny, které by sloužily jako úchytné body pro podpůrné vazy. Žádný z návrhů také nevysvětluje, proč má hřeben takový tvar, proč by ostatní lambeosauridi měli mít hřebeny, které vypadají mnohem jinak, ale plní podobnou funkci, jak se hadrosauridi bez hřebene nebo s pevným hřebenem obešli bez takových schopností nebo proč měli někteří hadrosauridi pevné hřebeny. Tyto úvahy ovlivňují zejména hypotézy založené na zvýšení schopností systémů, které jsou u zvířete již přítomny, jako jsou hypotézy o solné žláze a čichu, a naznačují, že tyto funkce nebyly primárními funkcemi hřebene. Práce na nosní dutině lambeosaurinů navíc ukazují, že čichové nervy a odpovídající smyslová tkáň byly z velké části mimo část nosních průduchů v hřebeni, takže rozšíření hřebene mělo jen málo společného s čichem.
Sociální funkce
Obnovení života Parasaurolophus walkeri s hypotetickou kožní řasou a pruhy na hřebeni.
Na místo toho se jako funkce (funkce) hřebene začaly více podporovat sociální a fyziologické funkce se zaměřením na vizuální a sluchovou identifikaci a komunikaci. Jako velký objekt má hřeben jasnou hodnotu jako vizuální signál a odlišuje toto zvíře od jeho současníků. Velké rozměry očních důlků hadrosauridů a přítomnost sklerotických kroužků v očích naznačují ostrý zrak a denní návyky, což je důkaz, že zrak byl pro tato zvířata důležitý. Pokud by se, jak je běžně znázorňováno, kožní řasa táhla od hřebene až na krk nebo záda, byl by navrhovaný vizuální projev mnohem okázalejší. Jak naznačují jiné lebky lambeosaurinů, hřeben parasaurolofů pravděpodobně umožňoval jak druhovou identifikaci (například oddělení od korytosaura nebo lambeosaura), tak určení mezi samci a samicemi na základě tvaru a velikosti.
Zvuková funkce
Počítačový model nosních průchodů v hřebeni P. tubicen.
Vnější vzhled hřebene však neodpovídá složité vnitřní anatomii nosních průchodů, což naznačuje jinou funkci zohledňující využití vnitřního prostoru. Carl Wiman jako první v roce 1931 navrhl, že průchody plní funkci sluchové signalizace, podobně jako krkavice; Hopson a David B. Weishampel se k této myšlence vrátili v 70. a 80. letech 20. století. Hopson zjistil, že existují anatomické důkazy, že hadrosauridi měli silný sluch. Existuje přinejmenším jeden příklad, u příbuzného korythosaura, kde je na místě štíhlý stapes (plazí ušní kost), což v kombinaci s velkým prostorem pro ušní bubínek naznačuje citlivé střední ucho. Kromě toho je lagena hadrosaurida protáhlá jako u krokodýla, což naznačuje, že sluchová část vnitřního ucha byla dobře vyvinutá. Weishampel předpokládal, že P. walkeri byl schopen produkovat frekvence 48 až 240 Hz a P. cyrtocristatus (interpretovaný jako juvenilní hřebenová forma) 75 až 375 Hz. Na základě podobnosti vnitřního ucha hadrosauridů s vnitřním uchem krokodýlů také navrhl, že dospělí hadrosauridi byli citliví na vysoké frekvence, jaké mohli produkovat jejich potomci. Podle Weishampela to odpovídá komunikaci rodičů a potomků.
Počítačové modelování dobře zachovalého exempláře P. tubicen se složitějšími dýchacími cestami než u P. walkeri umožnilo rekonstruovat možný zvuk, který jeho hřeben vydával. Hlavní dráha rezonuje na frekvenci kolem 30 Hz, ale komplikovaná anatomie dutin způsobuje ve zvuku špičky a údolí.
Chladicí funkce
Velká plocha a vaskularizace hřebene také naznačuje termoregulační funkci. P. E. Wheeler poprvé navrhl toto využití v roce 1978 jako způsob, jak udržet mozek v chladu. Přibližně ve stejné době navrhli termoregulaci také Teresa Maryańska a Osmólska a další zájem projevili Sullivan a Williamson. Diskuse Davida Evanse z roku 2006 o funkci hřebene lambeosaurů byla této myšlence nakloněna, přinejmenším jako počátečnímu faktoru evoluce rozšíření hřebene.
V médiích
Parasaurolophus je velmi populární dinosaurus. Objevil se ve filmu Disneyho Dinosaurus jako člen stáda. Několikrát se objevil také ve slavném dokumentu Souboj dinosaurů. Objevil se také v populárním televizním pořadu Prehistorický park, kde se stal kořistí velkých masožravců Deinosucha a Albertosaura. Několikrát se objevil ve filmech Jurský park, v prvním filmu jako člen stáda, ve druhém byl držen v zajetí lovci a ve třetím běžel spolu s korytosaurem. a postava Dweeba ve filmu Jsme zpátky! Příběh dinosaura“ je sám Parasaurolophus.
Parasaurolophus se objevuje také ve filmu Turok, jako učenlivý požírač rostlin, který normálně není škodlivý, ale může být agresivní, pokud je silně vyprovokován.
V Power Rangers je také Parasaurolophus zord: Dino Thunder a Power Rangers:
Parasaurolophus se krátce objeví na začátku filmu studia Disney Pixar Hodný dinosaurus.
Viz také
Parasaurolophus/Galerie
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, str. 229. Vydáno: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, kap: Parasaurolophus, strany 678-684. Vydáno: 1997, McFarland & Co, in Jefferson, North Carolina. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull a Wright, Hadrosaurian Dinosaurs of North America (Hadrosauričtí dinosauři Severní Ameriky), s. 209-213.
- Brett-Surman, Michael K. a Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, kap: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, strany 135-169. Vydáno, 2006, Indiana University Press, in Bloomington and Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; and Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) The Dinosauria, 2nd edition, chapter: Hadrosauridae, strana 438-463. Vydáno: 2004, University of California Press, v Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, volume 13, pages 1-32. (Parasaurolophus walkeri, nový rod a druh chocholatého trachodontního dinosaura). Vydáno: 1922, University of Toronto Studies, Geology Series.
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. a Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, ze série New Mexico Museum of Natural History and Science Bulletin, 15, strany 1-52. Vydáno v roce 2013. Vydáno: 1999, New Mexico Museum of Natural History and Science, v Albuqueque, Nové Mexiko.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, Journal of Paleontology, roč. 35, 3. vydání, na stranách 575-577.
- Gilmore, Charles W., On the genus Stephanosaurus, with the description of the type specimen of Lambeosaurus lambei, volume 38, issue 43, on pages 29-48, Parks. Vydáno: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. a Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) The Dinosauria, 1. vydání, kap: Hadrosauridae, strany 534-561. Vydáno: 1990, University of California Press v Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., and Reisz, Robert R. 2007. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta, z Journal of Vertebrate Paleontology, vol. 27 issue 2, na stranách 373-393.
- Godefroit, Pascal, Shuqin Zan; a Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China, from the Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, pages 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Published by New Mexico Museum of Natural History and Science, in Albuqueque, New Mexico. Pages 191-213.
- Liddell, Henry George a Robert Scott, 1980. A Greek-English Lexicon (Abridged Edition) Vyd: Oxford University Press ve Spojeném království. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. a Evans, David C., Currie, Phillip J. a Koppelhus, Eva (eds.). Dinosauří provinční park: A Spectacular Ancient Ecosystem Revealed (Velkolepý starověký ekosystém odhalen), kap: Ornithischian Dinosaurs. 2005, vyd: Indiana University Press, v Bloomingtonu. Pages 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; and Dupuis, Kevin, 2007. A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus. Journal of Vertebrate Paleontology, vol. 27, issue 3 pages 642-650.
- Currie, Phillip J; and Russell, Dale A. „Geographic and stratigraphic distribution of dinosaur remains“ in Dinosaur Provincial Park, str. 553.
- Weishampel, David B. (1990). „Dinosaur Distribution“, in The Dinosauria (1. vydání), s. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). „Dinosaur Distribution“, in The Dinosauria (2. vydání), s. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, z Nového Acta Regia Societas Scientarum Upsaliensis, série 4, svazek 7, číslo 5. (německy). Pages 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary (Cassellův latinský slovník), vydaný nakladatelstvím Cassell Ltd. (česky). 1979, vydání 5, v Londýně. ISBN 0-304-52257-0 Strana 883.
- Weishampel, David B. a Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) from Utah, z Journal of Paleontology, vol. 53, issue 6, pages 1422-1427.
- Eberth, David A. 2005. „The geology“, in Dinosaur Provincial Park, s. 54-82.
- Braman, Dennis R., a Koppelhus, Eva B. 2005. „Campanian palynomorphs“, in Dinosaur Provincial Park, s. 101-130.
- Russell, Dale A. An Odyssey in Time (Odysea v čase): Dinosaurs of North America, 1989. Nakladatelství: NorthWord Press, v Minocqua, Wisconsin. ISBN 1-55971-038-1. Strany 160-164.
- Bakker, Robert T. 1986. Dinosauří kacířství: Bakker: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, vydalo nakladatelství William Morrow v New Yorku. ISBN 0-8217-2859-8 Strana 194.
- 27.0 27.1 27.2 Norman David B. Ilustrovaná encyklopedie dinosaurů: An Original and Compelling Insight into Life in the Dinosaur Kingdom (Originální a přesvědčivý pohled na život v dinosauří říši), kap: Hadrosauridi II. 1985. Vydalo nakladatelství Crescent Books v New Yorku. Strany 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs, z časopisu Journal of Paleobiology, roč. 32, č. 1, str. 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs , z časopisu Journal of Paleobiology, ročník první, číslo první, strany 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology, z University of Chicago Press, na straně 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, z časopisu Orion (Mnichov), sv. 2. str. 525-532 (v německém jazyce).
- Sternberg, Charles M. 1935. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous [Kapucínští hadrosauři série Belly River ze svrchní křídy] z časopisu Canada Department of Mines Bulletin (Geological Series), svazek 77, číslo 52, na str. 1-37.
- Colbert, Edwin H. The Dinosaur Book (Kniha o dinosaurech): The Ruling Reptiles and their Relatives (Vládnoucí plazi a jejich příbuzní), vydalo v roce 1945 Americké přírodovědné muzeum, Man and Nature Publications, 14, v New Yorku. Strana 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas z časopisu Jarbuch Naturwissenschaften, ročník 12, číslo 36, na stranách 709-716. (Německy) 1924.
- Osmólska, Halszka, 1979. Nosní solné žlázy u dinosaurů, z časopisu Acta Palaeontologica Polonica, roč. 24, na stranách 205-215.
- 36.0 36.1 Ostrom, John H., 1962. The cranial crests of hadrosaurian dinosaurs, z časopisu Journal of Postilla, sv. 62, strany 1-29.
- Gish, Duane T., 1992. Dinosauři podle designu, vydalo nakladatelství Master Books, in Green Forest. ISBN 0-89051-165-9. Strana 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), z časopisu Journal of Paleobiology, roč. 7, č. 2, s. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, z Journal of Paleontology, roč. 55, č. 5, s. 1046-1057.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur, od Sandia National Laboratories (1997-12-05). Získáno 20. ledna 2009.
- Diegert, Carl F. a Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen from the Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. a Williamson, Thomas E., 1996. A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications, z Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborate CNS cooling structure in large dinosaurs Časopis Nature, roč. 275, na str. 441-443.
- Maryańska, Teresa a Osmólska, Halszka, v roce 1979. Aspects of hadrosaurian cranial anatomy, z časopisu Journal of Lethaia, sv. 12, na str. 265-273.
- Vědci pomocí digitální paleontologie vytvořili hlas dinosaura parasaurolofa; ze Sandia National Laboratories.
- Restaurace P. walkeri a P. cyrtocristatus, ve stejném měřítku, od Scotta Hartmana; na Skeletal Drawing.com.
- Stránka parasaurolofa na Enchanted Learning.com
.