

Parasaurolophus (wymawiane /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, powszechnie również /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; Saurolophus) to rodzaj dinozaura z rzędu ornitopodów z późnej kredy na terenie dzisiejszej Ameryki Północnej, około 76-73 mln lat temu. Był to roślinożerca, który chodził zarówno jako dwunóg, jak i czworonóg. Znane są trzy gatunki: P. walkeri (gatunek typu), P. tubicen i krótkoogoniasty P. cyrtocristatus. Szczątki znane są z Alberty w Kanadzie oraz z Nowego Meksyku i Utah w USA. Po raz pierwszy został opisany w 1922 roku przez Williama Parksa na podstawie czaszki i częściowego szkieletu z Alberty.
Parasaurolophus to hadrozauryd, należący do zróżnicowanej rodziny dinozaurów z kredy, znanych z zakresu dziwacznych ozdób głowy. Rodzaj ten znany jest z dużego, rozbudowanego grzebienia czaszkowego, który w największym przypadku tworzy długą, zakrzywioną rurę wystającą z czaszki ku górze i tyłowi. Charonosaurus z Chin, który mógł być jego najbliższym krewnym, miał podobną czaszkę i potencjalnie podobny grzebień. Grzebień był szeroko dyskutowany przez naukowców; panuje zgoda co do tego, że jego główne funkcje obejmowały wizualne rozpoznawanie gatunku i płci, rezonans akustyczny i termoregulację. Jest to jeden z rzadszych kaczodziobów, znany z zaledwie kilku dobrych okazów.
Parasaurolophus należy do najbardziej rozpoznawalnych dinozaurów i jest często przedstawiany w związanych z dinozaurami programach telewizyjnych, filmach, książkach, towarach i innych mediach. Alice Perry
Opis

Rozmiar Parasaurolophus walkeri w porównaniu z człowiekiem.
Jak w przypadku większości dinozaurów, szkielet Parasaurolophus jest niekompletnie poznany. Długość okazu typu P. walkeri szacowana jest na 9,5 metra (31 stóp). Jego czaszka ma około 1,6 metra (5,2 stopy) długości, włączając grzebień, podczas gdy czaszka P. tubicen ma ponad 2,0 metry (6,6 stopy) długości, co wskazuje na większe zwierzę. Jego waga szacowana jest na 2,5 tony (2,7 tony). Jego pojedyncza znana kończyna przednia była stosunkowo krótka jak na hadrozauryda, z krótką, ale szeroką łopatką. Kość udowa u P. walkeri mierzy 103 centymetry długości i jest solidna jak na swoją długość w porównaniu z innymi hadrozaurydami. Górne ramię i kości miednicy były również mocno zbudowane.
Podobnie jak inne hadrozaurydy, był w stanie chodzić na dwóch nogach lub czterech. Prawdopodobnie wolał zdobywać pożywienie na czterech nogach, ale biegał na dwóch. Kolce nerwowe kręgów były wysokie, co było powszechne u lambeozaurynów; najwyższe nad biodrami, zwiększały wysokość grzbietu. Znane są odciski skóry P. walkeri, ukazujące jednolite gruzełkowate łuski, ale bez większych struktur.
Parasaurolophus walkeri ze szczegółami skalowania.
Najbardziej zauważalną cechą był grzebień czaszki, który wystawał z tyłu głowy i składał się z kości przedszczękowej i nosowej. Okaz typu P. walkeri ma wcięcie w kolcach nerwowych w pobliżu miejsca, gdzie grzebień uderzyłby w plecy, ale może to być patologia specyficzna dla tego osobnika. William Parks, który nazwał ten rodzaj, wysunął hipotezę, że więzadło biegnące od grzebienia do wcięcia podtrzymuje głowę. Chociaż pomysł ten wydaje się mało prawdopodobny, Parasaurolophus jest czasem odtwarzany z płatem skóry od grzebienia do szyi.
Grzebień był pusty, z wyraźnymi rurkami prowadzącymi od każdego nozdrza do końca grzebienia przed odwróceniem kierunku i zmierzaniem z powrotem w dół grzebienia i do czaszki. Rurki były najprostsze u P. walkeri, a bardziej złożone u P. tubicen, gdzie niektóre rurki były ślepe, a inne spotykały się i rozdzielały. Podczas gdy P. walkeri i P. tubicen miały długie grzebienie o niewielkim zakrzywieniu, P. cyrtocristatus miał krótki grzebień o bardziej okrągłym profilu.
Klasyfikacja
Jak sama nazwa wskazuje, Parasaurolophus był początkowo uważany za blisko spokrewnionego z Saurolophus ze względu na powierzchownie podobny grzebień. Wkrótce jednak ponownie uznano go za członka podrodziny lambeozaurynów hadrozaurydów – Saurolophus jest hadrozaurynem. Zazwyczaj interpretuje się go jako osobny odłam lambeozaurydów, odrębny od Corythosaurus, Hypacrosaurus i Lambeosaurus z hełmem na głowie. Jego najbliższym znanym krewnym wydaje się być Charonosaurus, lambeozauryna o podobnej czaszce (ale jeszcze bez kompletnego grzebienia) z regionu Amur w północno-wschodnich Chinach, a oba te gatunki mogą tworzyć klad Parasaurolophini. P. cyrtocristatus, z krótkim, bardziej zaokrąglonym grzebieniem, może być najbardziej bazalnym z trzech znanych gatunków Parasaurolophus, może też reprezentować niedorosłe lub żeńskie okazy P. tubicen.
Historia
Poznanie i nazewnictwo
Odlew niekompletnego okazu typu Parasaurolophus walkeri w Warszawie.
Sznacząca „jaszczurka z bliskim grzebieniem” nazwa Parasaurolophus pochodzi od greckiego para/παρα „obok” lub „w pobliżu”, saurus/σαυρος „jaszczurka” i lophos/λοφος „grzebień”. Jest on oparty na ROM 768, czaszce i częściowym szkielecie, któremu brakuje większości ogona i tylnych nóg poniżej kolan, który został znaleziony przez ekipę terenową z Uniwersytetu w Toronto w 1920 r. w pobliżu Sand Creek wzdłuż rzeki Red Deer w Albercie w Kanadzie. Skały te znane są obecnie jako górnokredowa formacja Dinosaur Park z okresu kampanu. William Parks nazwał okaz P. walkeri na cześć Sir Byrona Edmunda Walkera, przewodniczącego Rady Powierników Royal Ontario Museum. Szczątki Parasaurolophus są rzadkie w Albercie, z tylko jedną inną częściową czaszką z (prawdopodobnie) formacji Dinosaur Park i trzema okazami z Dinosaur Park pozbawionymi czaszek, prawdopodobnie należącymi do tego rodzaju. W niektórych wykazach fauny pojawia się wzmianka o możliwym materiale P. walkeri z formacji Hell Creek w Montanie, jednostki skalnej z późnego maastrychtu. Występowanie to nie jest odnotowane przez Sullivana i Williamsona w ich przeglądzie rodzaju z 1999 roku i nie zostało szerzej omówione w innych miejscach.
W 1921 roku Charles H. Sternberg odzyskał częściową czaszkę (PMU.R1250) z tego, co obecnie znane jest jako nieco młodsza formacja Kirtland w hrabstwie San Juan w Nowym Meksyku. Okaz ten został wysłany do Uppsali w Szwecji, gdzie Carl Wiman opisał go jako drugi gatunek, P. tubicen, w 1931 roku. Epitet szczególny pochodzi od łacińskiego tǔbǐcěn „trębacz”. Druga, niemal kompletna czaszka P. tubicen (NMMNH P-25100) została znaleziona w Nowym Meksyku w 1995 roku. Korzystając z tomografii komputerowej tej czaszki, Robert Sullivan i Thomas Williamson w 1999 r. przedstawili monografię tego rodzaju, obejmującą aspekty anatomii i taksonomii, a także funkcje grzebienia. Williamson opublikował później niezależny przegląd szczątków, nie zgadzając się z wnioskami taksonomicznymi.
John Ostrom opisał kolejny dobry okaz (FMNH P27393) z Nowego Meksyku jako P. cyrtocristatus w 1961 roku. Zawiera on częściową czaszkę z krótkim, zaokrąglonym grzebieniem i dużą część szkieletu postczaszkowego z wyjątkiem stóp, szyi i części ogona. Jego specyficzna nazwa pochodzi od łacińskiego curtus „skrócony” i cristatus „grzebieniasty”. Okaz został znaleziony w górnej części formacji Fruitland lub, co bardziej prawdopodobne, w podstawie nadległej formacji Kirtland. Zasięg występowania tego gatunku został rozszerzony w 1979 roku, kiedy David B. Weishampel i James A. Jensen opisali częściową czaszkę z podobnym grzebieniem (BYU 2467) z kambaniastej formacji Kaiparowits w hrabstwie Garfield w stanie Utah. Od tego czasu w Utah znaleziono kolejną czaszkę z krótką/okrągłą morfologią grzebienia P. cyrtocristatus.
Gatunek
Odlew czaszki Parasaurolophus walkeri, Natural History Museum, London.
Parasaurolophus cyrtocristatus odlew czaszki, Field Museum of Natural History.
Typowy gatunek P. walkeri, z Alberty, znany jest z jednego okazu. Różni się od P. tubicen posiadaniem prostszych rur w grzebieniu, a od P. cyrtocristatus posiadaniem długiego, niezaokrąglonego grzebienia i dłuższego górnego ramienia niż przedramienia.
P. tubicen, z Nowego Meksyku, znany jest ze szczątków co najmniej trzech osobników. Jest to największy gatunek, z bardziej złożonymi przejściami powietrznymi w grzebieniu niż P. walkeri, i dłuższym, prostszym grzebieniem niż P. cyrtocristatus.
P. cyrtocristatus, z Nowego Meksyku i Utah, znany jest z trzech możliwych okazów. Jest to najmniejszy gatunek, z krótkim, zaokrąglonym grzebieniem. Jego niewielki rozmiar i forma grzebienia doprowadziły kilku naukowców do sugestii, że reprezentuje on młode osobniki lub samice P. tubicen, który pochodzi z mniej więcej tego samego czasu i z tej samej formacji w Nowym Meksyku. Jak zauważył Thomas Williamson, typowy materiał P. cyrtocristatus ma około 72% wielkości P. tubicen, co jest bliskie wielkości, przy której inne lambeozauryny zaczynają wykazywać ostateczny dymorfizm płciowy w grzebieniach (~70% wielkości dorosłego osobnika). Stanowisko to zostało odrzucone w ostatnich przeglądach dotyczących lambeozaurynów.
Paleobiologia
Paleoekologia
Gorgosaurus (po lewej) atakujący Parasaurolophus cyrtocristatus.
Parasaurolophus walkeri, z formacji Dinosaur Park, był członkiem zróżnicowanej i dobrze udokumentowanej fauny prehistorycznych zwierząt, w tym znanych dinozaurów, takich jak rogaty Centrosaurus, Styracosaurus i Chasmosaurus; kaczodzioby Prosaurolophus, Gryposaurus, Corythosaurus i Lambeosaurus; tyranozauryd Gorgosaurus; oraz opancerzone Edmontonia i Euoplocephalus. Był to rzadki składnik tej fauny. Formacja Dinosaur Park jest interpretowana jako nisko położone otoczenie rzek i terenów zalewowych, które z czasem stały się bardziej bagniste i podlegały wpływom morskim w miarę przesuwania się na zachód cieśniny Western Interior Seaway. Klimat był cieplejszy niż w dzisiejszej Albercie, bez przymrozków, ale z bardziej wilgotnymi i suchszymi porami roku. Drzewa iglaste były najwyraźniej dominującymi roślinami baldachimowymi, a pod nimi występowały paprocie, paprocie drzewiaste i rośliny okrytozalążkowe.
Gatunki z Nowego Meksyku dzieliły swoje środowisko z dużym zauropodem Alamosaurus, kaczodziobym Kritosaurus, rogatym Pentaceratops, opancerzonym Nodocephalosaurus, Saurornitholestes i obecnie nienazwanymi tyranozaurydami. Formacja Kirtland interpretowana jest jako rzeczne rozlewiska powstałe po cofnięciu się Zachodniego Morza Wewnętrznego. Dominującymi roślinami były drzewa iglaste, a rogate dinozaury chasmozaurynów były najwyraźniej bardziej powszechne niż hadrozaurydy.
Żywienie
Jako hadrozauryd, Parasaurolophus był dużym dwunożnym/czworonożnym roślinożercą, odżywiającym się roślinami za pomocą wyrafinowanej czaszki, która pozwalała na ruchy ścierające analogiczne do żucia. Jego zęby były nieustannie wymieniane i pakowane w baterie zębowe zawierające setki zębów, z których tylko garstka była w użyciu w danym momencie. Używał dzioba do zbierania materiału roślinnego, który był trzymany w szczękach przez organ przypominający policzek. Żerowanie odbywało się od ziemi aż do około 4 metrów (13 stóp) powyżej. Jak zauważył Bob Bakker, lambeozauryny mają węższe dzioby niż hadrozauryny, co sugeruje, że Parasaurolophus i jego krewni mogli odżywiać się bardziej selektywnie niż ich pozbawione grzebienia odpowiedniki o szerokich dziobach.
Grzebień czaszkowy
Wiele hipotez zostało wysuniętych co do funkcji, jakie pełnił grzebień czaszkowy Parasaurolophus, ale większość z nich została zdyskredytowana. Obecnie uważa się, że mógł on mieć kilka funkcji: wizualne wyświetlanie do identyfikacji gatunku i płci, wzmocnienie dźwięku do komunikacji i termoregulacji. Nie jest jasne, która z nich była najbardziej istotna w jakim czasie w ewolucji grzebienia i jego wewnętrznych przejść nosowych.
Różnice między gatunkami i stadiami rozwojowymi
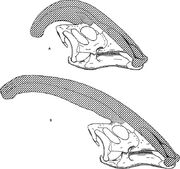
Diagram porównujący grzebienie narialne Parasaurolophus cyrtocristatus (a) i Parasaurolophus walkeri (b).
Podobnie jak u innych lambeozaurynów, uważa się, że grzebień czaszkowy Parasaurolophus zmieniał się wraz z wiekiem i był cechą dymorficzną płciowo u osobników dorosłych. James Hopson, jeden z pierwszych badaczy, którzy opisali grzebienie lambeozaurynów pod kątem takich rozróżnień, zasugerował, że P. cyrtocristatus, z małym grzebieniem, był żeńską formą P. tubicen. Thomas Williamson zasugerował, że była to forma młodociana. Żadna z hipotez nie zyskała powszechnej akceptacji. Ponieważ znanych jest tylko sześć dobrych czaszek i jedna młodociana mózgoczaszka, dodatkowy materiał pomoże wyjaśnić te potencjalne związki. Williamson zauważył, że w każdym razie młodociane Parasaurolophus prawdopodobnie miały małe, zaokrąglone grzebienie jak u P. cyrtocristatus, które prawdopodobnie rosły szybciej w miarę zbliżania się osobników do dojrzałości. Ostatnie ponowne badania młodocianej mózgoczaszki wcześniej przypisanej do Lambeosaurus, a obecnie do Parasaurolophus, dostarczyły dowodów na to, że mały rurkowaty grzebień występował u młodych osobników. Okaz ten zachowuje niewielkie, skierowane ku górze rozszczepienie kości czołowych, które było podobne, lecz mniejsze niż u dorosłych osobników; u dorosłych kości czołowe tworzyły platformę, która podtrzymywała podstawę grzebienia. Okaz ten wskazuje również, że wzrost grzebienia u Parasaurolophus i profil twarzy młodych osobników różnił się od modelu Corythosaurus-Hypacrosaurus-Lambeosaurus, częściowo dlatego, że grzebień Parasaurolophus pozbawiony jest cienkiego kostnego „grzebienia”, który tworzy górną część grzebienia pozostałych trzech lambeozaurynów.
Odrzucone hipotezy dotyczące funkcji
Wiele wczesnych sugestii skupiało się na przystosowaniach do wodnego trybu życia, podążając za hipotezą, że hadrozaurydy były amfibiami, co było powszechną linią myślenia aż do lat 60. XX wieku. Alfred Sherwood Romer zaproponował, że służyła ona jako rurka do nurkowania, Martin Wilfarth, że była to przystawka do ruchomej wyrostka robaczkowego używanego jako rurka do oddychania lub do zbierania pokarmu, Charles M. Sternberg, że służyła jako pułapka powietrzna, by utrzymać wodę z dala od płuc, a Ned Colbert, że służyła jako zbiornik powietrza przy dłuższych pobytach pod wodą.
Inne propozycje miały bardziej fizyczny charakter. Jak wspomniano powyżej, William Parks zasugerował, że był on połączony z kręgami za pomocą więzadeł lub mięśni, i pomagał w poruszaniu się i podtrzymywaniu głowy. Othenio Abel zaproponował, by służyła jako broń w walce między przedstawicielami tego samego gatunku, a Andrew Milner zasugerował, że mogła być używana jako deflektor liści, podobnie jak grzebień hełmu (zwany „casque”) sutenera. Jeszcze inne propozycje za główną funkcję uznawały obudowę wyspecjalizowanych narządów. Halszka Osmólska zasugerowała, że mieścił on gruczoły solne, a John Ostrom zasugerował, że mieścił rozszerzone obszary dla tkanki węchowej i znacznie ulepszony zmysł węchu lambeozaurynów, które nie miały oczywistych zdolności obronnych. Jedną z niezwykłych sugestii, wysuniętych przez kreacjonistę Duane’a Gisha, jest to, że grzebień mieścił gruczoły chemiczne, które pozwalały mu rzucać strumienie chemicznego „ognia” na wrogów, podobne do współczesnego chrząszcza bombardiera.
Większość z tych hipotez została zdyskredytowana lub odrzucona. Na przykład, nie ma otworu na końcu grzebienia dla funkcji nurkowania. Nie ma żadnych blizn po mięśniach dla trąbki i wątpliwe jest, czy zwierzę z dziobem potrzebowałoby jej. Jako proponowana śluza powietrzna, nie utrzymywałaby wody. Proponowany zbiornik powietrza byłby niewystarczający dla zwierzęcia wielkości Parasaurolophus. Inne hadrozaurydy miały duże głowy bez potrzeby posiadania dużych pustych grzebieni, które służyłyby jako punkty zaczepienia dla więzadeł podtrzymujących. Żadna z propozycji nie wyjaśnia też, dlaczego grzebień ma taki kształt, dlaczego inne lambeozauryny miałyby mieć grzebienie wyglądające inaczej, ale pełniące podobną funkcję, jak hadrozaurydy bez grzebienia lub z pełnym grzebieniem radziły sobie bez takich możliwości, ani dlaczego niektóre hadrozaurydy miały pełne grzebienie. Te rozważania szczególnie wpływają na hipotezy oparte na zwiększeniu możliwości systemów już obecnych u zwierzęcia, takie jak hipotezy gruczołu solnego i węchu, i wskazują, że nie były to pierwotne funkcje grzebienia. Dodatkowo, prace nad jamą nosową lambeozaurynów pokazują, że nerwy węchowe i odpowiadająca im tkanka czuciowa znajdowały się w dużej mierze poza częścią przejścia nosowego w grzebieniu, więc rozbudowa grzebienia miała niewiele wspólnego ze zmysłem węchu.
Funkcje społeczne
Przywrócenie do życia Parasaurolophus walkeri z hipotetyczną falbanką skórną i paskami na grzebieniu.
W zamian funkcje społeczne i fizjologiczne stały się bardziej wspierane jako funkcja(e) grzebienia, skupiając się na wizualnej i słuchowej identyfikacji i komunikacji. Jako duży obiekt, grzebień ma wyraźną wartość jako sygnał wizualny i odróżnia to zwierzę od jemu współczesnych. Duże rozmiary gałek ocznych hadrozaurydów i obecność pierścieni sklerotycznych w oczach sugerują ostre widzenie i dzienne zwyczaje, co świadczy o tym, że wzrok był dla tych zwierząt ważny. Gdyby, jak to jest powszechnie przedstawiane, falbanka skórna rozciągała się od grzebienia do szyi lub grzbietu, proponowany pokaz wizualny byłby znacznie bardziej efektowny. Jak sugerują inne czaszki lambeozaurynów, grzebień Parasaurolophus prawdopodobnie umożliwiał zarówno identyfikację gatunku (np. oddzielenie go od Corythosaurus czy Lambeosaurus), jak i rozróżnienie samców i samic na podstawie kształtu i rozmiaru.
Funkcja dźwiękowa
Komputerowy model przejść nosowych w grzebieniu P. tubicen.
Zewnętrzny wygląd grzebienia nie odpowiada jednak złożonej anatomii wewnętrznej przejść nosowych, co sugeruje inną funkcję odpowiedzialną za wykorzystanie przestrzeni wewnętrznej. Carl Wiman był pierwszym, który zaproponował w 1931 roku, że przejścia służyły słuchowej funkcji sygnalizacyjnej, jak crumhorn; Hopson i David B. Weishampel powrócili do tego pomysłu w latach 70. i 80. Hopson stwierdził, że istnieją anatomiczne dowody na to, że hadrozaurydy miały silny słuch. U spokrewnionego z nim koryntozaura istnieje co najmniej jeden przykład smukłego strzemiączka (reptiliańska kość słuchowa) na swoim miejscu, co w połączeniu z dużą przestrzenią na błonę bębenkową sugeruje istnienie wrażliwego ucha środkowego. Co więcej, lagena hadrozaurydów jest wydłużona jak u krokodyli, co wskazuje, że część słuchowa ucha wewnętrznego była dobrze rozwinięta. Weishampel zasugerował, że P. walkeri był w stanie wytwarzać częstotliwości od 48 do 240 Hz, a P. cyrtocristatus (interpretowany jako młodociana forma grzebieniasta) od 75 do 375 Hz. Opierając się na podobieństwie uszu wewnętrznych hadrozaurydów do uszu krokodyli, zaproponował on również, że dorosłe hadrozaurydy były wrażliwe na wysokie częstotliwości, takie jakie może wytwarzać ich potomstwo. Według Weishampela jest to zgodne z porozumiewaniem się rodziców i potomstwa.
Modelowanie komputerowe dobrze zachowanego okazu P. tubicen, posiadającego bardziej złożone kanały powietrzne niż te u P. walkeri, pozwoliło na odtworzenie możliwego dźwięku, jaki wydawał jego grzebień. Główna ścieżka rezonuje przy około 30 Hz, ale skomplikowana anatomia zatoki powoduje szczyty i doliny w dźwięku.
Funkcja chłodząca
Duża powierzchnia i unaczynienie grzebienia również sugeruje funkcję termoregulacyjną. P.E. Wheeler pierwszy zasugerował to użycie w 1978 roku jako sposób na utrzymanie mózgu w chłodzie. Teresa Maryańska i Osmólska również zaproponowały termoregulację mniej więcej w tym samym czasie, a Sullivan i Williamson podjęli dalsze zainteresowanie. Dyskusja Davida Evansa z 2006 roku na temat funkcji grzebienia lambeozaurynów sprzyjała temu pomysłowi, przynajmniej jako początkowemu czynnikowi ewolucji ekspansji grzebienia.
W mediach
Parasaurolophus to bardzo popularny dinozaur. Wystąpił w filmie Disney’s Dinosaur jako członek stada. Wystąpił również w słynnym filmie dokumentalnym Clash of the Dinosaurs. Pojawił się także w popularnym programie telewizyjnym Prehistoric Park, gdzie stał się ofiarą dużych drapieżników Deinosuchus i Albertosaurus. Kilka razy pojawił się w filmach z serii Jurassic Park, w pierwszym filmie jako członek stada, w drugim w niewoli myśliwych, a w trzecim biegał razem z koryntozaurem. oraz postać Dweeba w filmie „We’re Back! A Dinosaur’s Story”, jest Parasaurolophus sam.
Parasaurolophus pojawia się również w Turok, jako potulny zjadacz roślin, który normalnie nie jest szkodliwy, ale może być agresywny, jeśli poważnie sprowokowany.
Jest również Parasaurolophus zord w Power Rangers: Dino Thunder i Power Rangers: Dino Charge.
Parasaurolophus pojawia się na krótko na początku filmu Disney Pixar The Good Dinosaur.
See Also
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, strona 229. Published: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Chapter: Parasaurolophus, strony 678-684. Published: 1997, McFarland & Co, in Jefferson, North Carolina. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull i Wright, Hadrosaurian Dinosaurs of North America, s. 209-213.
- Brett-Surman, Michael K. and Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, rozdział: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, strony 135-169. Published, 2006, Indiana University Press, in Bloomington and Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; and Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) The Dinosauria, 2nd edition, chapter: Hadrosauridae, strony 438-463. Wydane: 2004, University of California Press, w Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, tom 13, strony 1-32. Published: 1922, University of Toronto Studies, Geology Series.
- 7,0 7,1 7,2 7,3 7,4 7,5 7,6 7,7 Sullivan, Robert M. and Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, z serii New Mexico Museum of Natural History and Science Bulletin, tom 15, strony 1-52. Opublikowane: 1999, New Mexico Museum of Natural History and Science, w Albuqueque, Nowy Meksyk.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaurian from the Cretaceous of New Mexico, Journal of Paleontology, Volume 35, 3rd issue, na stronach 575-577.
- Gilmore, Charles W., On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, volume 38, issue 43, pages 29-48, Parks. Published: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. and Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) The Dinosauria, 1st edition, Chapter: Hadrosauridae, strony 534-561. Wydane: 1990, University of California Press w Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., and Reisz, Robert R. 2007. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta, z Journal of Vertebrate Paleontology, vol. 27 issue 2, na stronach 373-393.
- Godefroit, Pascal, Shuqin Zan; and Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China, z Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, na stronach 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Published by New Mexico Museum of Natural History and Science, in Albuqueque, New Mexico. Strony 191-213.
- Liddell, Henry George i Robert Scott, 1980. A Greek-English Lexicon (Abridged Edition) Opublikowano: Oxford University Press w Wielkiej Brytanii. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. and Evans, David C., Currie, Phillip J., and Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, Chapter: Ornithischian Dinosaurs. 2005, Opublikowano: Indiana University Press, w Bloomington. Pages 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; and Dupuis, Kevin, 2007. A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments ontogeny crest in the genus. Journal of Vertebrate Paleontology, vol. 27, issue 3 pages 642-650.
- Currie, Phillip J; and Russell, Dale A. „Geographic and stratigraphic distribution of dinosaur remains” w Dinosaur Provincial Park, str. 553.
- Weishampel, David B. (1990). „Rozmieszczenie dinozaurów”, w The Dinosauria (1st), s. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). „Rozmieszczenie dinozaurów”, w: The Dinosauria (2nd), s. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, z Nova Acta Regia Societas Scientarum Upsaliensis, series 4, vol. 7, issue 5. (niemiecki). Strony 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, opublikowane przez Cassell Ltd., wydanie 5, w Londynie. 1979, wydanie 5, w Londynie. ISBN 0-304-52257-0 Strona 883.
- Weishampel, David B. and Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) from Utah, z Journal of Paleontology, vol. 53, issue 6, pages 1422-1427.
- Eberth, David A. 2005. „The geology”, w Dinosaur Provincial Park, s. 54-82.
- Braman, Dennis R., and Koppelhus, Eva B. 2005. „Campanian palynomorphs”, w Dinosaur Provincial Park, pp. 101-130.
- Russell, Dale A. An Odyssey in Time: Dinosaurs of North America, 1989. Wydawca: NorthWord Press, w Minocqua, Wisconsin. ISBN 1-55971-038-1 Strony 160-164.
- Bakker, Robert T. 1986. The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, opublikowane przez William Morrow, w Nowym Jorku. ISBN 0-8217-2859-8 Strona 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom (Oryginalny i fascynujący wgląd w życie w królestwie dinozaurów), rozdział: Hadrozaurydy II. 1985. Wydane przez Crescent Books, w Nowym Jorku. Strony 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs, z Journal of Paleobiology, vol. 32, issue 1, Pages 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs , z Journal of Paleobiology, volume one, issue one, strony 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology, z University of Chicago Press, na stronie 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, z Journal of Orion (Munich), vol. 2. s. 525-532 (język niemiecki).
- Sternberg, Charles M. 1935. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous from the Journal of the Canada Department of Mines Bulletin (Geological Series), volume 77, issue 52, on pages 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, opublikowana w 1945 roku przez American Museum of Natural History, Man and Nature Publications, 14, w Nowym Jorku. Strona 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas z czasopisma Jarbuch Naturwissenschaften, tom 12, numer 36, na stronach 709-716. (niemiecki) 1924.
- Osmólska, Halszka, 1979. Gruczoły solne nosowe u dinozaurów, z czasopisma Acta Palaeontologica Polonica, vol. 24, na stronach 205-215.
- 36.0 36.1 Ostrom, John H., 1962. The cranial crests of hadrosaurian dinosaurs, z czasopisma Journal of Postilla, vol. 62, strony 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, wydane przez Master Books, w Green Forest. ISBN 0-89051-165-9 Strona 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), z Journal of Paleobiology, vol. 7, issue 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, z Journal of Paleontology, vol. 55, issue 5, pp. 1046-1057.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur, by the Sandia National Laboratories (1997-12-05). Retrieved on January 20th, 2009.
- Diegert, Carl F. i Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen z Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. and Williamson, Thomas E., 1996. A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications, z Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborate CNS chłodzenia struktury w dużych dinozaurów Journal of Nature, vol. 275, na str. 441-443.
- Maryańska, Teresa i Osmólska, Halszka, w 1979. Aspekty anatomii czaszki hadrozaurów, z Journal of Lethaia, vol. 12, na s. 265-273.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur; z Sandia National Laboratories.
- Restorations of P. walkeri and P. cyrtocristatus, to the same scale, by Scott Hartman; at Skeletal Drawing.com.
- Parasaurolophus page on Enchanted Learning.com
.