

Parasaurolophus (udtales /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, almindeligvis også /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; betyder “nær kammøllet øgle” med henvisning til Saurolophus) er en slægt af ornithopode-dinosaurer fra den sene kridttid i det nuværende Nordamerika for ca. 76-73 millioner år siden. Det var en planteæder, der gik både som tobenet og firbenet. Der er anerkendt tre arter: Der findes tre arter: P. walkeri (typearten), P. tubicen og den kortskæggede P. cyrtocristatus. Rester er kendt fra Alberta, Canada, og New Mexico og Utah, USA. Den blev første gang beskrevet i 1922 af William Parks fra et kranie og et delvist skelet i Alberta.
Parasaurolophus er en hadrosaurid, der er en del af en forskelligartet familie af dinosaurer fra kridttiden, der er kendt for deres række bizarre hovedudsmykninger. Denne slægt er kendt for sin store, udførlige kraniekrone, som i sin største form danner et langt buet rør, der rager opad og bagud fra kraniet. Charonosaurus fra Kina, som kan have været dens nærmeste slægtning, havde et lignende kranium og muligvis en lignende kam. Forskerne har diskuteret kammen meget; der er enighed om, at de vigtigste funktioner omfatter visuel genkendelse af både art og køn, akustisk resonans og termoregulering. Den er en af de sjældnere andeskaller, idet den kun er kendt fra en håndfuld gode eksemplarer.
Parasaurolophus er blandt de mest velkendte dinosaurer og er ofte portrætteret i dinosaurierelaterede tv-serier, film, bøger, merchandise og andre medier. Alice Perry
Beskrivelse

Størrelsen af Parasaurolophus walkeri sammenlignet med et menneske.
Som det er tilfældet med de fleste dinosaurer, er skelettet af Parasaurolophus ufuldstændigt kendt. Længden af typeeksemplaret af P. walkeri er anslået til 9,5 meter (31 ft). Dens kranium er ca. 1,6 meter langt, inklusive kam, mens typeskallen af P. tubicen er over 2,0 meter lang, hvilket tyder på et større dyr. Dens vægt er anslået til 2,5 tons (2,7 tons). Dens eneste kendte forben var relativt kort for en hadrosaurid, med et kort, men bredt skulderblad. Lårbenet måler 103 centimeter (3,38 ft) hos P. walkeri og er robust for sin længde sammenlignet med andre hadrosaurider. Overarm- og bækkenknoglerne var også kraftigt bygget.
Som andre hadrosaurider var den i stand til at gå på enten to eller fire ben. Den foretrak sandsynligvis at søge efter føde på fire ben, men løb på to. Hvirvlernes neuralrygge var høje, som det var almindeligt hos lambeosauriner; de var højest over hofterne og øgede ryggens højde. Der kendes hudaftryk fra P. walkeri, der viser ensartede tuberkellignende skæl, men ingen større strukturer.
Parasaurolophus walkeri med skældetalje.
Det mest iøjnefaldende træk var kraniekronen, der stak ud fra baghovedet og bestod af forkammen og næseknogler. Typeeksemplaret af P. walkeri har et hak i neuralryggene nær det sted, hvor kammen ville ramme ryggen, men dette kan være en patologi, der er særegent for dette individ. William Parks, som navngav slægten, opstillede den hypotese, at et ledbånd løb fra kammen til hakket for at støtte hovedet. Selv om ideen virker usandsynlig, er Parasaurolophus nogle gange restaureret med en hudlap fra kammen til halsen.
Kammen var hul, med tydelige rør, der førte fra hvert næsebor til kammerets ende, før de vendte retning og gik tilbage ned ad kammen og ind i kraniet. Rørene var de enkleste hos P. walkeri og mere komplekse hos P. tubicen, hvor nogle rør var blinde og andre mødtes og adskilte sig. Mens P. walkeri og P. tubicen havde lange kamme med kun svag krumning, havde P. cyrtocristatus en kort kam med en mere cirkulær profil.
Klassifikation
Som navnet antyder, blev Parasaurolophus oprindeligt anset for at være nært beslægtet med Saurolophus på grund af dens overfladisk set ens kamme. Den blev dog hurtigt omvurderet som et medlem af den lambeosauriske underfamilie af hadrosaurider – Saurolophus er en hadrosaurin. Den tolkes normalt som en særskilt udløber af lambeosaurinerne, der adskiller sig fra de hjelmskjoldede Corythosaurus, Hypacrosaurus og Lambeosaurus. Dens nærmeste kendte slægtning synes at være Charonosaurus, en lambeosaurine med et lignende kranium (men endnu ikke en komplet kam) fra Amur-regionen i det nordøstlige Kina, og de to kan danne en klade Parasaurolophini. P. cyrtocristatus, med sin korte, rundere kam, kan være den mest basale af de tre kendte Parasaurolophus-arter, eller den kan repræsentere subadulte eller hunlige eksemplarer af P. tubicen.
Historie
Afstøbning af det ufuldstændige typeeksemplar af Parasaurolophus walkeri i Warszawa.
Navnet Parasaurolophus er afledt af græsk para/παρα “ved siden af” eller “nær”, saurus/σαυρος “firben” og lophos/λοφος “krone”. Den er baseret på ROM 768, et kranium og et delvist skelet, der mangler det meste af halen og bagbenene under knæene, som blev fundet af et felthold fra University of Toronto i 1920 nær Sand Creek langs Red Deer River i Alberta, Canada. Disse bjergarter er nu kendt som Dinosaur Park-formationen fra Campanian-alderen i den øvre kridttid fra Dinosaur Park. William Parks navngav eksemplaret P. walkeri til ære for Sir Byron Edmund Walker, formand for bestyrelsen for Royal Ontario Museum of the Royal Ontario Museum. Parasaurolophus-rester er sjældne i Alberta, med kun ét andet delvist kranium fra (sandsynligvis) Dinosaur Park-formationen og tre Dinosaur Park-eksemplarer uden kranier, der muligvis tilhører slægten. I nogle faunalister er der en omtale af muligt P. walkeri-materiale i Hell Creek Formationen i Montana, en bjergartsenhed af sen maastrichtiansk alder. Denne forekomst er ikke nævnt af Sullivan og Williamson i deres gennemgang af slægten fra 1999 og er ikke blevet uddybet yderligere andre steder.
I 1921 genfandt Charles H. Sternberg et delvist kranium (PMU.R1250) fra det, der nu er kendt som den lidt yngre Kirtland Formation i San Juan County, New Mexico. Dette eksemplar blev sendt til Uppsala i Sverige, hvor Carl Wiman beskrev det som en anden art, P. tubicen, i 1931. Det specifikke epithet er afledt af det latinske tǔbǐcěn “trompeter”. Et andet, næsten komplet P. tubicen-kranie (NMMNH P-25100) blev fundet i New Mexico i 1995. Ved hjælp af computertomografi af dette kranium gav Robert Sullivan og Thomas Williamson slægten en monografisk behandling i 1999, der dækkede aspekter af dens anatomi og taxonomi samt kammefunktionerne. Williamson offentliggjorde senere en uafhængig gennemgang af resterne og var uenig i de taksonomiske konklusioner.
John Ostrom beskrev et andet godt eksemplar (FMNH P27393) fra New Mexico som P. cyrtocristatus i 1961. Det omfatter et delvist kranium med en kort, afrundet kam og meget af det postkraniale skelet bortset fra fødderne, halsen og dele af halen. Dens specifikke navn er afledt af det latinske curtus “forkortet” og cristatus “krone”. Eksemplaret blev fundet i enten toppen af Fruitland-formationen eller, mere sandsynligt, i bunden af den overliggende Kirtland-formation. Artens udbredelsesområde blev udvidet i 1979, da David B. Weishampel og James A. Jensen beskrev et delvist kranie med en lignende krone (BYU 2467) fra Kaiparowits-formationen fra Campanian-alderen i Garfield County, Utah. Siden da er endnu et kranie blevet fundet i Utah med den korte/runde P. cyrtocristatus-kammemorfologi.
Species
Parasaurolophus walkeri kranieafstøbning, Natural History Museum, London.
Parasaurolophus cyrtocristatus kranieafstøbning, Field Museum of Natural History.
Typearten P. walkeri, fra Alberta, er kendt fra et enkelt eksemplar. Den adskiller sig fra P. tubicen ved at have enklere rør i kammen og fra P. cyrtocristatus ved at have en lang, uafrundet kam og en længere overarm end underarm.
P. tubicen, fra New Mexico, er kendt fra resterne af mindst tre individer. Det er den største art, med mere komplekse luftpassager i kammen end P. walkeri, og en længere, mere lige kam end P. cyrtocristatus.
P. cyrtocristatus, fra New Mexico og Utah, er kendt fra tre mulige eksemplarer. Det er den mindste art, med en kort afrundet kam. Dens lille størrelse og kamformen har fået flere forskere til at foreslå, at den repræsenterer unge individer eller hunner af P. tubicen, som er fra nogenlunde samme tid og fra den samme formation i New Mexico. Som Thomas Williamson har bemærket, er typematerialet af P. cyrtocristatus ca. 72 % af størrelsen på P. tubicen, hvilket er tæt på den størrelse, hvor andre lambeosauriner fortolkes til at begynde at vise definitiv kønsdimorphisme i deres kamme (~70 % af voksenstørrelse). Denne holdning er blevet afvist i nyere anmeldelser af lambeosauriner.
Paleobiology
Paleoecology
Gorgosaurus (til venstre) angriber en Parasaurolophus cyrtocristatus.
Parasaurolophus walkeri fra Dinosaur Park-formationen var medlem af en mangfoldig og veldokumenteret fauna af forhistoriske dyr, herunder velkendte dinosaurer som den hornede Centrosaurus, Styracosaurus og Chasmosaurus; andeskaller Prosaurolophus, Gryposaurus, Corythosaurus og Lambeosaurus; tyrannosaurid Gorgosaurus; og pansrede Edmontonia og Euoplocephalus. Den var en sjælden bestanddel af denne fauna. Dinosaur Park Formationen fortolkes som en lavreliefindstilling af floder og flodsletter, der blev mere sumpede og påvirket af marine forhold med tiden, efterhånden som den vestlige indre havvej trængte vestpå. Klimaet var varmere end i det nuværende Alberta, uden frost, men med vådere og mere tørre årstider. Nåletræer var tilsyneladende de dominerende planter i kronetaget med et underlag af bregner, træfarne og angiospermer.
De new mexicanske arter delte deres miljø med den store sauropod Alamosaurus, Kritosaurus med andeskralde, Pentaceratops med horn, Nodocephalosaurus med panser, Saurornitholestes og tyrannosaurider, der endnu ikke er navngivet. Kirtland-formationen tolkes som flodflodsletter, der opstod efter tilbagetrækning af den vestlige indre søvej. Nåletræer var de dominerende planter, og chasmosauriske horndinosaurer var tilsyneladende mere almindelige end hadrosaurider.
Spisning
Som hadrosaurid var Parasaurolophus en stor tobenet/kvadrobenet planteæder, der spiste planter med et sofistikeret kranium, der tillod en malende bevægelse svarende til tyggebevægelse. Dens tænder blev løbende udskiftet og pakket ind i tandbatterier, der indeholdt hundredvis af tænder, hvoraf kun en relativ håndfuld var i brug på et hvilket som helst tidspunkt. Den brugte sit næb til at beskære plantemateriale, som blev holdt fast i kæberne af et kindlignende organ. Den kunne æde fra jorden og op til ca. 4 meter over jorden. Som Bob Bakker har bemærket, har lambeosauriner smalere næb end hadrosauriner, hvilket indebærer, at Parasaurolophus og dens slægtninge kunne æde mere selektivt end deres brednæbbede, kamløse modstykker.
Kraniekrone
Mange hypoteser er blevet fremsat om, hvilke funktioner kraniekronen hos Parasaurolophus udførte, men de fleste er blevet miskrediteret. Man mener nu, at den kan have haft flere funktioner: visuel visning til identifikation af art og køn, lydforstærkning til kommunikation og termoregulering. Det er ikke klart, hvilken der var mest betydningsfuld på hvilke tidspunkter i udviklingen af kammen og dens indre næsegange.
Differencer mellem arter og vækststadier
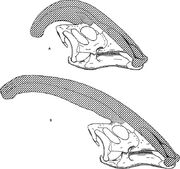
Diagram, der sammenligner narialkammene hos Parasaurolophus cyrtocristatus (a) og Parasaurolophus walkeri (b).
Som for andre lambeosauriner antages det, at kraniekammen hos Parasaurolophus ændrede sig med alderen og var et kønsdimorft kendetegn hos voksne. James Hopson, en af de første forskere, der beskrev lambeosaurinernes kamme i form af sådanne kendetegn, foreslog, at P. cyrtocristatus, med sin lille kam, var hunformen af P. tubicen. Thomas Williamson foreslog, at det var den juvenile form. Ingen af disse hypoteser blev bredt accepteret. Da der kun kendes seks gode kranier og en juvenil hjernekasse, vil yderligere materiale hjælpe med at klarlægge disse potentielle relationer. Williamson bemærkede, at under alle omstændigheder havde juvenile Parasaurolophus sandsynligvis små, afrundede kamme ligesom P. cyrtocristatus, der sandsynligvis voksede hurtigere, efterhånden som individerne nærmede sig modenhed. En nyere genundersøgelse af en juvenil hjernekasse, der tidligere blev tildelt Lambeosaurus og nu tildelt Parasaurolophus, giver beviser for, at en lille rørformet kam var til stede hos juvenile dyr. Dette eksemplar har bevaret en lille opadgående udblæsning af frontalknoglerne, der lignede, men var mindre end det, der ses i voksne eksemplarer; hos voksne dannede frontalknoglerne en platform, der støttede kamens basis. Dette eksemplar viser også, at kamens vækst hos Parasaurolophus og ansigtsprofilen hos unge individer adskilte sig fra Corythosaurus-Hypacrosaurus-Lambeosaurus-modellen, til dels fordi kammen hos Parasaurolophus mangler den tynde knoglelignende “coxcomb”, der udgør den øverste del af kammen hos de tre andre lambeosauriner.
Forkastede hypoteser om funktion
Mange tidlige forslag fokuserede på tilpasninger til en akvatisk livsstil, i forlængelse af hypotesen om, at hadrosaurider var amfibiske, en almindelig tankegang indtil 1960’erne. Således foreslog Alfred Sherwood Romer, at den tjente som snorkel, Martin Wilfarth, at den var en fastgørelse til en bevægelig proboscis, der blev brugt som åndedrætsrør eller til at samle føde, Charles M. Sternberg, at den tjente som luftfang for at holde vand ude af lungerne, og Ned Colbert, at den tjente som luftreservoir ved længerevarende ophold under vandet.
Andre forslag var af mere fysisk karakter. Som nævnt ovenfor foreslog William Parks, at den var forbundet til ryghvirvlerne med ledbånd eller muskler og hjalp med at bevæge og støtte hovedet. Othenio Abel foreslog, at den blev brugt som et våben i kamp mellem medlemmer af samme art, og Andrew Milner foreslog, at den kunne bruges som en løvafviser, ligesom kasuarens hjelmkam (kaldet en “casque”). I andre forslag var den vigtigste funktion at huse specialiserede organer. Halszka Osmólska foreslog, at den rummede saltkirtler, og John Ostrom foreslog, at den rummede udvidede områder for olfaktorisk væv og en meget forbedret lugtesans hos lambeosaurinerne, som ikke havde nogen åbenlyse forsvarsevner. Et usædvanligt forslag, fremsat af kreationisten Duane Gish, er, at kammen husede kemiske kirtler, der gjorde det muligt for den at kaste stråler af kemisk “ild” mod fjender, svarende til den moderne bombardierbille.
De fleste af disse hypoteser er blevet miskrediteret eller afvist. For eksempel er der ikke noget hul for enden af kammen til en snorkelfunktion. Der er ingen muskelar til en proboscis, og det er tvivlsomt, at et dyr med et næb ville have brug for et næb. Som en foreslået luftsluse ville den ikke have holdt vand ude. Det foreslåede luftreservoir ville ikke have været tilstrækkeligt til et dyr på størrelse med Parasaurolophus. Andre hadrosaurider havde store hoveder uden at have brug for store hule kamme, der kunne tjene som fastgørelsespunkter for støttebånd. Ingen af forslagene forklarer heller ikke, hvorfor kammen har en sådan form, hvorfor andre lambeosauriner skulle have kamme, der ser meget anderledes ud, men som udfører en lignende funktion, hvordan kamløse hadrosaurider eller hadrosaurider med faste kamme klarede sig uden sådanne funktioner, eller hvorfor nogle hadrosaurider havde faste kamme. Disse overvejelser påvirker især hypoteser, der er baseret på en forøgelse af kapaciteten af systemer, der allerede findes i dyret, såsom saltkirtel- og lugthypoteserne, og tyder på, at disse ikke var crestens primære funktioner. Desuden viser arbejdet med næsehulen hos lambeosaurinerne, at olfaktoriske nerver og tilsvarende sensorisk væv stort set lå uden for den del af næsepassagerne i kammen, så udvidelsen af kammen havde kun lidt at gøre med lugtesansen.
Sociale funktioner
Livsrestaurering af Parasaurolophus walkeri med hypotetisk hudkrave og striber på kammen.
I stedet er sociale og fysiologiske funktioner blevet mere understøttet som funktion(er) af kammen, med fokus på visuel og auditiv identifikation og kommunikation. Som et stort objekt har kammen en klar værdi som et visuelt signal og adskiller dette dyr fra sine samtidige artsfæller. Den store størrelse af hadrosauridernes øjenhuler og tilstedeværelsen af sklerotiske ringe i øjnene antyder et skarpt syn og døgnvaner, hvilket beviser, at synet var vigtigt for disse dyr. Hvis der, som det ofte er illustreret, var en hudkrølle, der strakte sig fra kammen til nakken eller ryggen, ville det foreslåede visuelle display have været meget mere prangende. Som det antydes af andre lambeosauriske kranier, tillod Parasaurolophus’ kam sandsynligvis både artsidentifikation (som f.eks. adskillelse fra Corythosaurus eller Lambeosaurus) og bestemmelse mellem hanner og hunner, baseret på form og størrelse.
Lydfunktion
Computermodel af næsepassagerne i kammen hos P. tubicen.
Den ydre fremtoning af kammen svarer imidlertid ikke til den komplekse indre anatomi af næsepassagerne, hvilket tyder på en anden funktion, der redegjorde for brugen af det indre rum. Carl Wiman var den første, der i 1931 foreslog, at passagerne tjente en auditiv signalfunktion, ligesom et krummehorn; Hopson og David B. Weishampel genoptog denne idé i 1970’erne og 1980’erne. Hopson fandt, at der er anatomiske beviser for, at hadrosaurider havde en stærk hørelse. Der er mindst ét eksempel, hos den beslægtede Corythosaurus, på en slank stigbøjle (reptiløreknogle) på plads, hvilket sammen med et stort rum til trommehinden antyder et følsomt mellemøre. Desuden er hadrosauridens lagena langstrakt som en krokodilles, hvilket indikerer, at den auditive del af det indre øre var veludviklet. Weishampel foreslog, at P. walkeri var i stand til at producere frekvenser på 48 til 240 Hz, og P. cyrtocristatus (tolket som en juvenil kamform) 75 til 375 Hz. Baseret på ligheden mellem hadrosauridernes indre øre og krokodillernes, foreslog han også, at voksne hadrosaurider var følsomme over for høje frekvenser, som deres afkom kunne producere. Ifølge Weishampel er dette i overensstemmelse med, at forældre og afkom kunne kommunikere.
Computermodellering af et velbevaret eksemplar af P. tubicen, med mere komplekse luftkanaler end hos P. walkeri, har gjort det muligt at rekonstruere den mulige lyd, som dens kam producerede. Hovedvejen resonerer ved omkring 30 Hz, men den komplicerede sinusanatomi forårsager toppe og dale i lyden.
Kølende funktion
Den store overflade og vaskularisering af kammen tyder også på en termoregulerende funktion. P.E. Wheeler foreslog først denne anvendelse i 1978 som en måde at holde hjernen kølig på. Teresa Maryańska og Osmólska foreslog også termoregulering på omtrent samme tidspunkt, og Sullivan og Williamson viste yderligere interesse. David Evans’ diskussion i 2006 om lambeosaurernes kamfunktioner var positiv over for ideen, i hvert fald som en indledende faktor for udviklingen af kamudvidelse.
I medierne
Parasaurolophus er en meget populær dinosaur. Den var med i filmen Disney’s Dinosaur som et flokmedlem. Den optrådte også flere gange i den berømte dokumentarfilm Clash of the Dinosaurs. Den optrådte også i det populære tv-show Prehistoric Park, hvor den blev bytte for de store kødædere Deinosuchus og Albertosaurus. Den optrådte et par gange i Jurassic Park-filmene, som medlem af en flok i den første film, hvorefter den blev holdt fanget af jægere i den anden film og løb sammen med Corythosaurus i den tredje film. og karakteren Dweeb i “We’re Back! A Dinosaur’s Story” er selv en Parasaurolophus.
Parasaurolophus optræder også i Turok, som en føjelig planteæder, der normalt ikke er skadelig, men som kan være aggressiv, hvis den bliver voldsomt provokeret.
Der findes også en Parasaurolophus-zord i Power Rangers: Dino Thunder og Power Rangers: Dino Charge.
Parasaurolophus optræder kortvarigt i begyndelsen af Disney Pixar-filmen The Good Dinosaur.
Se også
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, side 229. Udgivet: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Chapter: Parasaurolophus, side 678-684. Udgivet: 1997, McFarland & Co, i Jefferson, North Carolina. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull and Wright, Hadrosaurian Dinosaurs of North America, pp. 209-213.
- Brett-Surman, Michael K. og Wagner, Jonathan R. Carpenter, Kenneth (red.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, kapitel: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, kapitel: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, side 135-169. Udgivet, 2006, Indiana University Press, i Bloomington og Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; og Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; og Dodson, Peter (red.) The Dinosauria, 2. udgave, kapitel: The Dinosauria, 2nd edition, chapter: Hadrosauridae, side 438-463. Udgivet: 2004, University of California Press, i Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, volume 13, pages 1-32. Udgivet: 1922, University of Toronto Studies, Geology Series.
- 7.0 7.1 7.1 7.2 7.3 7.4 7.4 7.5 7.6 7.7 Sullivan, Robert M. og Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, fra serien New Mexico Museum of Natural History and Science Bulletin, 15, pages 1-52. Udgivet: 1999, New Mexico Museum of Natural History and Science, i Albuqueque, New Mexico.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, Journal of Paleontology, Volume 35, 3rd issue, on pages 575-577.
- Gilmore, Charles W., On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, volume 38, issue 43, pages 29-48, Parks. Udgivet: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. og Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) The Dinosauria, 1. udgave, kapitel: Hadrosauridae, side 534-561. Udgivet: 1990, University of California Press i Berkeley. ISBN 0-520-0672727-4
- 11.0 11.1 11.2 11.3 Evans, David C., og Reisz, Robert R. 2007. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta, fra Journal of Vertebrate Paleontology, vol. 27 issue 2, på siderne 373-393.
- Godefroit, Pascal, Shuqin Zan; og Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaur fra det sene Maastrichtium i det nordøstlige Kina, fra Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, pages 875-882.
- 13.0 13.1 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Published by New Mexico Museum of Natural History and Science, in Albuquequeque, New Mexico. Siderne 191-213.
- Liddell, Henry George og Robert Scott, 1980. A Greek-English Lexicon (forkortet udgave) Udgivet: Oxford University Press i Det Forenede Kongerige. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. and Evans, David C., Currie, Phillip J., and Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, kapitel: Ornithischian Dinosaurs. 2005, Udgivet: Indiana University Press, i Bloomington. Siderne 312-348, ISBN 0-253-3459595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; og Dupuis, Kevin, 2007. A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus. Journal of Vertebrate Paleontology, vol. 27, issue 3 pages 642-650.
- Currie, Phillip J; og Russell, Dale A. “Geographic and stratigraphic distribution of dinosaur remains” i Dinosaur Provincial Park, s. 553.
- Weishampel, David B. (1990). “Dinosaur Distribution”, i The Dinosauria (1st), s. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; og Noto, Christopher R. (2004). “Dinosaur Distribution”, i The Dinosauria (2nd), pp. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, fra Nova Acta Regia Societas Scientarum Upsaliensis, serie 4, vol. 7, issue 5. (tysk). Siderne 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, udgivet af Cassell Ltd. 1979, 5. udgave, i London. ISBN 0-304-52257-0 Side 883.
- Weishampel, David B. og Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) fra Utah, fra Journal of Paleontology, vol. 53, issue 6, pages 1422-1427.
- Eberth, David A. 2005. “The geology”, i Dinosaur Provincial Park, s. 54-82.
- Braman, Dennis R., og Koppelhus, Eva B. 2005. “Campanian palynomorphs”, i Dinosaur Provincial Park, pp. 101-130.
- Russell, Dale A. An Odyssey in Time: Dinosaurs of North America, 1989. Forlag: Forlaget: NorthWord Press, i Minocqua, Wisconsin. ISBN 1-55971-038-1 Siderne 160-164.
- Bakker, Robert T. 1986. The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, udgivet af William Morrow, i New York. ISBN 0-8217-2859-8 Side 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom, kapitel: Hadrosaurider II. 1985. Udgivet af Crescent Books, i New York. Siderne 122-127. ISBN 0-517-468905
- 28.0 28.1 28.1 28.2 28.2 28.3 28.4 Evans, David C., 2006. Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs, fra Journal of Paleobiology, vol. 32, issue 1, Pages 109-125.
- 29.0 29.1 29.1 29.2 29.3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs , from the Journal of Paleobiology, volume one, issue one, pages 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology , fra University of Chicago Press, på side 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, fra Journal of Orion (München), bd. 2. s. 525-532 (tysk sprog).
- Sternberg, Charles M. 1935. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous from the Journal of the Canada Department of Mines Bulletin (Geological Series), volume 77, issue 52, on pages 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, udgivet i 1945 af American Museum of Natural History, Man and Nature Publications, 14, i New York. Side 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas fra Journal of Jarbuch Naturwissenschaften, bind 12, nummer 36, på siderne 709-716. (tysk) 1924.
- Osmólska, Halszka, 1979. Nasale saltkirtler hos dinosaurer, fra Journal of Acta Palaeontologica Polonica, vol. 24, på siderne 205-215.
- 36.0 36.1 Ostrom, John H., 1962. The cranial crests of hadrosaurian dinosaurs, fra Journal of Postilla, vol. 62, pages 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, udgivet af Master Books, i Green Forest. ISBN 0-89051-165-9 Side 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), fra Journal of Paleobiology, vol. 7, issue 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, from the Journal of Paleontology, vol. 55, issue 5, pp. 1046-1057.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur, fra Sandia National Laboratories (1997-12-05). Hentet den 20. januar 2009.
- Diegert, Carl F. og Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen from the Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. og Williamson, Thomas E., 1996. A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications, fra Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborate CNS cooling structure in large dinosaurs Journal of Nature, vol. 275, on pp. 441-443.
- Maryańska, Teresa og Osmólska, Halszka, i 1979. Aspects of hadrosaurian cranial anatomy, fra Journal of Lethaia, vol. 12, på s. 265-273.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur; fra Sandia National Laboratories.
- Restaureringer af P. walkeri og P. cyrtocristatus, i samme målestok, af Scott Hartman; på Skeletal Drawing.com.
- Parasaurolophus side på Enchanted Learning.com