

Parasaurolophus (ausgesprochen /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, umgangssprachlich auch /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; in Anlehnung an Saurolophus) ist eine Gattung ornithopoder Dinosaurier aus der späten Kreidezeit des heutigen Nordamerikas vor etwa 76-73 Millionen Jahren. Er war ein Pflanzenfresser, der sich sowohl als Zwei- als auch als Vierbeiner fortbewegte. Es sind drei Arten bekannt: P. walkeri (die Typusart), P. tubicen, und der kurzköpfige P. cyrtocristatus. Überreste sind aus Alberta, Kanada, und New Mexico und Utah, USA, bekannt. Er wurde erstmals 1922 von William Parks anhand eines Schädels und eines Teilskeletts in Alberta beschrieben.
Parasaurolophus ist ein Hadrosaurier, der zu einer vielfältigen Familie kreidezeitlicher Dinosaurier gehört, die für ihre bizarren Kopfverzierungen bekannt sind. Diese Gattung ist für ihren großen, kunstvollen Schädelkamm bekannt, der in seiner größten Ausprägung eine lange, gebogene Röhre bildet, die vom Schädel nach oben und hinten absteht. Charonosaurus aus China, der möglicherweise sein nächster Verwandter war, hatte einen ähnlichen Schädel und möglicherweise eine ähnliche Schädeldecke. Der Kamm ist unter Wissenschaftlern viel diskutiert worden; man ist sich einig, dass die wichtigsten Funktionen die visuelle Erkennung von Art und Geschlecht, akustische Resonanz und Thermoregulation waren. Er gehört zu den selteneren Entenschnäbeln und ist nur von einer Handvoll guter Exemplare bekannt.
Parasaurolophus gehört zu den bekanntesten Dinosauriern und wird häufig in Fernsehsendungen, Filmen, Büchern, Merchandising-Artikeln und anderen Medien mit Bezug zu Dinosauriern dargestellt. Alice Perry
Beschreibung

Die Größe von Parasaurolophus walkeri im Vergleich zu einem Menschen.
Wie bei den meisten Dinosauriern ist das Skelett von Parasaurolophus nur unvollständig bekannt. Die Länge des Typusexemplars von P. walkeri wird auf 9,5 Meter (31 ft) geschätzt. Sein Schädel ist etwa 1,6 m lang, einschließlich des Kammes, während der Typusschädel von P. tubicen über 2,0 m lang ist, was auf ein größeres Tier hindeutet. Sein Gewicht wird auf 2,5 Tonnen (2,7 t) geschätzt. Sein einziges bekanntes Vorderbein war für einen Hadrosauriden relativ kurz, mit einem kurzen, aber breiten Schulterblatt. Der Oberschenkelknochen misst bei P. walkeri 103 Zentimeter und ist im Vergleich zu anderen Hadrosauriden für seine Länge sehr robust. Auch die Oberarm- und Beckenknochen waren kräftig gebaut.
Wie andere Hadrosauriden konnte er entweder auf zwei oder vier Beinen laufen. Er zog es wahrscheinlich vor, auf vier Beinen nach Nahrung zu suchen, lief aber auf zwei Beinen. Die Neuralstacheln der Wirbel waren hoch, wie es bei Lambeosauriern üblich war; sie waren am höchsten über den Hüften und vergrößerten die Höhe des Rückens. Von P. walkeri sind Hautabdrücke bekannt, die einheitliche, tuberkelartige Schuppen, aber keine größeren Strukturen zeigen.
Parasaurolophus walkeri with scalation detail.
Das auffälligste Merkmal war der Schädelkamm, der von der Rückseite des Kopfes abstand und aus dem Prämaxilla und den Nasenknochen bestand. Das Typusexemplar von P. walkeri hat eine Kerbe in den Neuralstacheln in der Nähe der Stelle, an der der Kamm auf den Rücken stößt, aber das könnte eine Pathologie sein, die diesem Individuum eigen ist. William Parks, der die Gattung benannte, stellte die Hypothese auf, dass ein Band vom Kamm zur Kerbe verläuft, um den Kopf zu stützen. Obwohl dieser Gedanke unwahrscheinlich erscheint, wird Parasaurolophus manchmal mit einem Hautlappen vom Kamm bis zum Hals wiederhergestellt.
Der Kamm war hohl, mit deutlichen Röhren, die von jedem Nasenloch zum Ende des Kammes führten, bevor sie die Richtung umkehrten und den Kamm hinunter in den Schädel zurückführten. Die Röhren waren bei P. walkeri am einfachsten und bei P. tubicen komplexer, da einige Röhren blind waren und andere sich trafen und trennten. Während P. walkeri und P. tubicen lange Kämme mit nur leichter Krümmung hatten, hatte P. cyrtocristatus einen kurzen Kamm mit einem eher kreisförmigen Profil.
Klassifizierung
Wie der Name schon sagt, wurde Parasaurolophus aufgrund seines oberflächlich ähnlichen Kammes zunächst für eng mit Saurolophus verwandt gehalten. Er wurde jedoch bald als Mitglied der Lambeosaurier-Unterfamilie der Hadrosaurier eingestuft – Saurolophus ist ein Hadrosaurier. Er wird gewöhnlich als ein separater Ableger der Lambeosaurier interpretiert, der sich von den behelmten Corythosaurus, Hypacrosaurus und Lambeosaurus unterscheidet. Sein engster bekannter Verwandter scheint Charonosaurus zu sein, ein Lambeosaurier mit einem ähnlichen Schädel (aber noch keinem vollständigen Kamm) aus der Amur-Region im Nordosten Chinas, und die beiden könnten eine Klade Parasaurolophini bilden. P. cyrtocristatus, mit seinem kurzen, runderen Kamm, könnte die basalste der drei bekannten Parasaurolophus-Arten sein, oder es könnte sich um subadulte oder weibliche Exemplare von P. tubicen handeln.
Geschichte
Entdeckung und Namensgebung
Abdruck des unvollständigen Typusexemplars von Parasaurolophus walkeri in Warszawa.
Der Name Parasaurolophus bedeutet „nahe Schopfeidechse“ und leitet sich vom griechischen para/παρα „neben“ oder „nahe“, saurus/σαυρος „Eidechse“ und lophos/λοφος „Schopf“ ab. Er basiert auf ROM 768, einem Schädel und Teilskelett, bei dem der größte Teil des Schwanzes und die Hinterbeine unterhalb der Knie fehlten, die 1920 von einem Forscherteam der Universität Toronto in der Nähe des Sand Creek am Red Deer River in Alberta, Kanada, gefunden wurden. Dieses Gestein ist heute als Dinosaur Park Formation aus der oberen Kreidezeit (Campanium) bekannt. William Parks nannte das Exemplar zu Ehren von Sir Byron Edmund Walker, dem Vorsitzenden des Kuratoriums des Royal Ontario Museum, P. walkeri. Überreste von Parasaurolophus sind in Alberta selten, es gibt nur einen weiteren Teilschädel (wahrscheinlich) aus der Dinosaur Park Formation und drei Exemplare aus der Dinosaur Park Formation ohne Schädel, die möglicherweise zu dieser Gattung gehören. In einigen Faunenlisten wird mögliches Material von P. walkeri in der Hell Creek Formation von Montana, einer Gesteinseinheit aus dem späten Maastrichtium, erwähnt. Dieses Vorkommen wird von Sullivan und Williamson in ihrer 1999 erschienenen Übersichtsarbeit über die Gattung nicht erwähnt und wurde auch an anderer Stelle nicht weiter ausgeführt.
Im Jahr 1921 entdeckte Charles H. Sternberg einen Teilschädel (PMU.R1250) aus der heute als etwas jünger bekannten Kirtland-Formation in San Juan County, New Mexico. Dieses Exemplar wurde nach Uppsala, Schweden, geschickt, wo Carl Wiman es 1931 als eine zweite Art, P. tubicen, beschrieb. Das spezifische Epitheton ist vom lateinischen tǔbǐcěn „Trompeter“ abgeleitet. Ein zweiter, fast vollständiger Schädel von P. tubicen (NMMNH P-25100) wurde 1995 in New Mexico gefunden. Anhand der Computertomographie dieses Schädels behandelten Robert Sullivan und Thomas Williamson 1999 die Gattung monographisch, wobei sie Aspekte der Anatomie und Taxonomie sowie die Funktionen des Kammes berücksichtigten. Williamson veröffentlichte später einen unabhängigen Bericht über die Überreste, in dem er den taxonomischen Schlussfolgerungen nicht zustimmte.
John Ostrom beschrieb 1961 ein weiteres gutes Exemplar (FMNH P27393) aus New Mexico als P. cyrtocristatus. Es enthält einen Teilschädel mit einem kurzen, abgerundeten Kamm und einen Großteil des postkranialen Skeletts mit Ausnahme der Füße, des Halses und Teilen des Schwanzes. Der spezifische Name leitet sich vom lateinischen curtus „verkürzt“ und cristatus „gekrönt“ ab. Das Exemplar wurde entweder im oberen Teil der Fruitland-Formation oder, was wahrscheinlicher ist, in der Basis der darüber liegenden Kirtland-Formation gefunden. Das Verbreitungsgebiet dieser Art wurde 1979 erweitert, als David B. Weishampel und James A. Jensen einen Teilschädel mit einem ähnlichen Kamm (BYU 2467) aus der Kaiparowits-Formation des Garfield County, Utah, aus dem Campanium beschrieben. Seitdem wurde in Utah ein weiterer Schädel mit der kurzen/runden Kammmorphologie von P. cyrtocristatus gefunden.
Spezies
Parasaurolophus walkeri Schädelabguss, Natural History Museum, London.
Parasaurolophus cyrtocristatus Schädelabguss, Field Museum of Natural History.
Die Typusart P. walkeri, aus Alberta, ist von einem einzigen Exemplar bekannt. Sie unterscheidet sich von P. tubicen durch einfachere Röhren in ihrem Kamm und von P. cyrtocristatus durch einen langen, unrunden Kamm und einen längeren Oberarm als Unterarm.
P. tubicen, aus New Mexico, ist von den Überresten von mindestens drei Individuen bekannt. Es ist die größte Art, mit komplexeren Luftkanälen in ihrem Kamm als P. walkeri und einem längeren, geraderen Kamm als P. cyrtocristatus.
P. cyrtocristatus, aus New Mexico und Utah, ist von drei möglichen Exemplaren bekannt. Es ist die kleinste Art mit einem kurzen, abgerundeten Scheitel. Ihre geringe Größe und die Form ihres Kammes haben mehrere Wissenschaftler zu der Vermutung veranlasst, dass es sich um Jungtiere oder Weibchen von P. tubicen handelt, die ungefähr aus der gleichen Zeit und aus der gleichen Formation in New Mexico stammt. Wie Thomas Williamson feststellte, ist das Typusmaterial von P. cyrtocristatus etwa 72 % so groß wie das von P. tubicen, was in etwa der Größe entspricht, bei der andere Lambeosaurier beginnen, einen definitiven Geschlechtsdimorphismus in ihren Kämmen zu zeigen (~70 % der Erwachsenengröße). Diese Position wurde in neueren Übersichten über Lambeosaurinen abgelehnt.
Paläobiologie
Paläoökologie
Gorgosaurus (links) greift einen Parasaurolophus cyrtocristatus an.
Parasaurolophus walkeri, aus der Dinosaurierpark-Formation, war ein Mitglied einer vielfältigen und gut dokumentierten Fauna prähistorischer Tiere, darunter bekannte Dinosaurier wie der gehörnte Centrosaurus, Styracosaurus und Chasmosaurus; die Entenschnäbel Prosaurolophus, Gryposaurus, Corythosaurus und Lambeosaurus, den Tyrannosaurus Gorgosaurus sowie die gepanzerten Edmontonia und Euoplocephalus. Er war ein seltener Bestandteil dieser Fauna. Die Dinosaur Park Formation wird als ein Flachrelief mit Flüssen und Überschwemmungsgebieten interpretiert, das im Laufe der Zeit sumpfiger wurde und unter dem Einfluss mariner Bedingungen stand, als der westliche innere Seeweg nach Westen vordrang. Das Klima war wärmer als im heutigen Alberta, ohne Frost, aber mit feuchteren und trockeneren Jahreszeiten. Nadelbäume waren offenbar die vorherrschenden Baumkronenpflanzen mit einem Unterwuchs aus Farnen, Baumfarnen und Angiospermen.
Die Arten aus New Mexico teilten sich ihre Umgebung mit dem großen Sauropoden Alamosaurus, dem Entenschnabel Kritosaurus, dem gehörnten Pentaceratops, dem gepanzerten Nodocephalosaurus, Saurornitholestes und derzeit unbenannten Tyrannosauriden. Die Kirtland-Formation wird als Flussüberschwemmungsgebiet interpretiert, das nach dem Rückzug des westlichen inneren Seewegs entstand. Koniferen waren die vorherrschenden Pflanzen, und chasmosaurische Hornsaurier waren offenbar häufiger als Hadrosaurier.
Fütterung
Als Hadrosaurier war Parasaurolophus ein großer zweibeiniger/vierbeiniger Pflanzenfresser, der Pflanzen mit einem hochentwickelten Schädel fraß, der eine dem Kauen ähnliche Mahlbewegung ermöglichte. Seine Zähne wurden ständig ersetzt und in Zahnbatterien gepackt, die Hunderte von Zähnen enthielten, von denen immer nur eine relative Handvoll in Gebrauch war. Er benutzte seinen Schnabel, um Pflanzen zu ernten, die von einem wangenartigen Organ im Kiefer gehalten wurden. Die Fütterung erfolgte vom Boden aus bis zu einer Höhe von etwa 4 m (13 ft). Wie Bob Bakker feststellte, haben Lambeosaurier schmalere Schnäbel als Hadrosaurier, was darauf hindeutet, dass Parasaurolophus und seine Verwandten selektiver fressen konnten als ihre breitschnäbligen, kammlosen Artgenossen.
Schädelkamm
Viele Hypothesen wurden aufgestellt, welche Funktionen der Schädelkamm von Parasaurolophus erfüllte, aber die meisten wurden widerlegt. Heute geht man davon aus, dass er mehrere Funktionen hatte: Er diente der visuellen Darstellung zur Identifizierung von Art und Geschlecht, der Schallverstärkung zur Kommunikation und der Wärmeregulierung. Es ist nicht klar, welche Funktion zu welchem Zeitpunkt in der Entwicklung des Nasenrückens und der inneren Nasengänge am wichtigsten war.
Unterschiede zwischen Arten und Wachstumsstadien
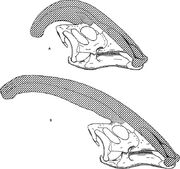
Diagramm zum Vergleich der Nasenrücken von Parasaurolophus cyrtocristatus (a) und Parasaurolophus walkeri (b).
Wie bei anderen Lambeosauriern wird angenommen, dass sich die Schädeldecke von Parasaurolophus mit dem Alter veränderte und bei Erwachsenen ein geschlechtsdimorphes Merkmal war. James Hopson, einer der ersten Forscher, der die Schädeldecke von Lambeosauriern anhand solcher Unterscheidungen beschrieb, schlug vor, dass P. cyrtocristatus mit seiner kleinen Schädeldecke die weibliche Form von P. tubicen war. Thomas Williamson schlug vor, es handele sich um die juvenile Form. Keine der beiden Hypothesen wurde allgemein akzeptiert. Da nur sechs gute Schädel und ein juveniler Hirnschädel bekannt sind, wird zusätzliches Material dazu beitragen, diese möglichen Beziehungen zu klären. Williamson merkte an, dass juvenile Parasaurolophus in jedem Fall wahrscheinlich kleine, abgerundete Kämme wie P. cyrtocristatus hatten, die wahrscheinlich schneller wuchsen, je näher die Individuen der Reife kamen. Die jüngste Untersuchung eines juvenilen Gehirns, das früher Lambeosaurus und jetzt Parasaurolophus zugeordnet wurde, liefert den Beweis, dass ein kleiner röhrenförmiger Kamm bei juvenilen Tieren vorhanden war. Bei diesem Exemplar ist eine kleine, nach oben gerichtete Aufweitung der Stirnknochen erhalten, die ähnlich, aber kleiner war als bei erwachsenen Exemplaren; bei Erwachsenen bildeten die Stirnknochen eine Plattform, die die Basis des Kammes stützte. Dieses Exemplar deutet auch darauf hin, dass das Wachstum des Kammes bei Parasaurolophus und das Gesichtsprofil der jugendlichen Individuen vom Corythosaurus-Hypacrosaurus-Lambeosaurus-Modell abweicht, zum Teil deshalb, weil dem Kamm von Parasaurolophus der dünne knöcherne „Coxcomb“ fehlt, der den oberen Teil des Kammes der anderen drei Lambeosaurier ausmacht.
Abgelehnte Hypothesen über die Funktion
Viele frühe Vorschläge konzentrierten sich auf Anpassungen an eine aquatische Lebensweise und folgten der Hypothese, dass Hadrosauriden amphibisch waren, eine bis in die 1960er Jahre gängige Denkweise. So schlug Alfred Sherwood Romer vor, dass es als Schnorchel diente, Martin Wilfarth, dass es ein Aufsatz für einen beweglichen Rüssel war, der als Atemschlauch oder zum Nahrungssammeln verwendet wurde, Charles M. Sternberg, dass es als Luftsperre diente, um Wasser aus der Lunge fernzuhalten, und Ned Colbert, dass es als Luftreservoir für längere Aufenthalte unter Wasser diente.
Andere Vorschläge waren eher physischer Natur. Wie bereits erwähnt, schlug William Parks vor, dass er durch Bänder oder Muskeln mit den Wirbeln verbunden war und bei der Bewegung und Stützung des Kopfes half. Othenio Abel schlug vor, dass er im Kampf zwischen Artgenossen als Waffe eingesetzt wurde, und Andrew Milner schlug vor, dass er als Laubabweiser dienen könnte, ähnlich wie der Helmkamm (genannt „casque“) des Kasuars. Andere Vorschläge sahen in der Unterbringung spezialisierter Organe die Hauptfunktion. Halszka Osmólska schlug vor, dass er Salzdrüsen beherbergte, und John Ostrom schlug vor, dass er erweiterte Bereiche für das Geruchsgewebe und einen viel besseren Geruchssinn der Lambeosaurier beherbergte, die keine offensichtlichen Verteidigungsfähigkeiten hatten. Ein ungewöhnlicher Vorschlag, der vom Kreationisten Duane Gish gemacht wurde, ist, dass der Kamm chemische Drüsen beherbergte, die es ihm ermöglichten, chemische „Feuerstrahlen“ auf Feinde zu schleudern, ähnlich wie der moderne Bombardierkäfer.
Die meisten dieser Hypothesen wurden diskreditiert oder verworfen. Zum Beispiel gibt es kein Loch am Ende des Kammes für eine Schnorchelfunktion. Es gibt keine Muskelnarben für einen Rüssel, und es ist zweifelhaft, dass ein Tier mit einem Schnabel einen solchen benötigen würde. Als vorgeschlagene Luftschleuse hätte sie kein Wasser zurückgehalten. Das vorgeschlagene Luftreservoir wäre für ein Tier von der Größe von Parasaurolophus nicht ausreichend gewesen. Andere Hadrosauriden hatten große Köpfe, ohne große hohle Kämme zu benötigen, die als Befestigungspunkte für stützende Bänder dienten. Außerdem erklärt keiner der Vorschläge, warum der Kamm eine solche Form hat, warum andere Lambeosaurier Kämme haben sollten, die ganz anders aussehen, aber eine ähnliche Funktion erfüllen, wie Hadrosauriden ohne Kamm oder mit festem Kamm ohne solche Fähigkeiten auskamen, oder warum einige Hadrosauriden feste Kämme hatten. Diese Überlegungen wirken sich insbesondere auf Hypothesen aus, die auf der Erweiterung der Fähigkeiten von bereits im Tier vorhandenen Systemen beruhen, wie z. B. die Salzdrüsen- und Geruchshypothese, und deuten darauf hin, dass dies nicht die primären Funktionen des Kammes waren. Darüber hinaus zeigen Arbeiten an der Nasenhöhle von Lambeosauriern, dass Geruchsnerven und entsprechendes sensorisches Gewebe größtenteils außerhalb des Teils der Nasengänge im Kamm lagen, so dass die Erweiterung des Kammes wenig mit dem Geruchssinn zu tun hatte.
Soziale Funktionen
Lebensrestauration von Parasaurolophus walkeri mit hypothetischer Hautkrause und Streifen auf dem Kamm.
Soziale und physiologische Funktionen wurden stattdessen stärker als Funktion(en) des Kammes unterstützt, wobei der Schwerpunkt auf visueller und auditiver Identifikation und Kommunikation lag. Als großes Objekt hat der Kamm einen klaren Wert als visuelles Signal und hebt dieses Tier von seinen Zeitgenossen ab. Die Größe der Augenhöhlen der Hadrosaurier und das Vorhandensein von sklerotischen Ringen in den Augen deuten auf ein scharfes Sehvermögen und Tagesgewohnheiten hin, was beweist, dass das Sehvermögen für diese Tiere wichtig war. Wenn sich, wie häufig dargestellt, eine Hautkrause vom Kamm bis zum Nacken oder Rücken erstreckte, wäre die vorgeschlagene visuelle Darstellung viel auffälliger gewesen. Wie andere Lambeosaurier-Schädel nahelegen, ermöglichte der Kamm von Parasaurolophus wahrscheinlich sowohl die Artbestimmung (z. B. zur Unterscheidung von Corythosaurus oder Lambeosaurus) als auch die Unterscheidung zwischen Männchen und Weibchen anhand von Form und Größe.
Lautfunktion
Computermodell der Nasenkanäle im Kamm von P. tubicen.
Das äußere Erscheinungsbild des Kammes entspricht jedoch nicht der komplexen inneren Anatomie der Nasenkanäle, was auf eine andere Funktion schließen lässt, die für die Nutzung des inneren Raumes verantwortlich ist. Carl Wiman war der erste, der 1931 vorschlug, dass die Nasengänge wie ein Krummhorn eine auditive Signalfunktion haben; Hopson und David B. Weishampel griffen diese Idee in den 1970er und 1980er Jahren wieder auf. Hopson stellte fest, dass es anatomische Beweise dafür gibt, dass Hadrosauriden ein starkes Gehör hatten. Beim verwandten Corythosaurus gibt es mindestens ein Beispiel für einen schlanken Steigbügel (Reptilienohrknochen), der zusammen mit einem großen Raum für das Trommelfell auf ein empfindliches Mittelohr schließen lässt. Darüber hinaus ist die Laga der Hadrosaurier länglich wie die eines Krokodils, was darauf hindeutet, dass der auditive Teil des Innenohrs gut entwickelt war. Weishampel schlug vor, dass P. walkeri in der Lage war, Frequenzen von 48 bis 240 Hz zu erzeugen, und P. cyrtocristatus (interpretiert als juvenile Kammform) 75 bis 375 Hz. Aufgrund der Ähnlichkeit der Innenohren von Hadrosauriden mit denen von Krokodilen schlug er außerdem vor, dass erwachsene Hadrosauriden für hohe Frequenzen empfindlich waren, wie sie ihre Nachkommen erzeugen könnten. Weishampel zufolge stimmt dies mit der Kommunikation zwischen Eltern und Nachkommen überein.
Die Computermodellierung eines gut erhaltenen Exemplars von P. tubicen mit komplexeren Luftkanälen als die von P. walkeri ermöglichte die Rekonstruktion des möglichen Klangs, den sein Scheitel erzeugte. Der Hauptweg resoniert bei etwa 30 Hz, aber die komplizierte Sinusanatomie verursacht Spitzen und Täler im Klang.
Kühlfunktion
Die große Oberfläche und die Vaskularisierung des Scheitels lassen auch auf eine thermoregulatorische Funktion schließen. P.E. Wheeler schlug diese Funktion erstmals 1978 vor, um das Gehirn kühl zu halten. Teresa Maryańska und Osmólska schlugen etwa zur gleichen Zeit ebenfalls eine Thermoregulation vor, und Sullivan und Williamson zeigten weiteres Interesse. Die 2006 von David Evans geführte Diskussion über die Funktion des Lambeosaurierkamms begünstigte diese Idee, zumindest als anfänglicher Faktor für die Evolution der Kammexpansion.
In den Medien
Parasaurolophus ist ein sehr populärer Dinosaurier. Er war in dem Film Disney’s Dinosaurier als Herdenmitglied zu sehen. Er hatte auch mehrere Auftritte in dem berühmten Dokumentarfilm Clash of the Dinosaurs. Er trat auch in der beliebten Fernsehserie Prehistoric Park auf, wo er die Beute der großen Fleischfresser Deinosuchus und Albertosaurus wurde. Er hatte einige Auftritte in den Jurassic-Park-Filmen, als Herdenmitglied im ersten Film, dann als Gefangener von Jägern im zweiten und als Mitläufer von Corythosaurus im dritten. und die Figur Dweeb in „We’re Back! A Dinosaur’s Story“ ist selbst ein Parasaurolophus.
Der Parasaurolophus kommt auch in Turok vor, als gelehriger Pflanzenfresser, der normalerweise nicht schädlich ist, aber aggressiv werden kann, wenn er stark provoziert wird.
Es gibt auch einen Parasaurolophus-Zord in Power Rangers: Dino Thunder und Power Rangers: Dino Charge.
Parasaurolophus erscheint kurz am Anfang des Disney Pixar Films The Good Dinosaur.
See Also
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, Seite 229. Veröffentlicht: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Chapter: Parasaurolophus, Seiten 678-684. Veröffentlicht: 1997, McFarland & Co, in Jefferson, North Carolina. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull und Wright, Hadrosaurian Dinosaurs of North America, S. 209-213.
- Brett-Surman, Michael K. und Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, Kapitel: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, Seiten 135-169. Veröffentlicht, 2006, Indiana University Press, in Bloomington und Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; und Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; und Dodson, Peter (eds.) The Dinosauria, 2nd edition, chapter: Hadrosauridae, Seiten 438-463. Veröffentlicht: 2004, University of California Press, in Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, Band 13, Seiten 1-32. Veröffentlicht: 1922, University of Toronto Studies, Geology Series.
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. and Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, from the series New Mexico Museum of Natural History and Science Bulletin, 15, pages 1-52. Veröffentlicht: 1999, New Mexico Museum of Natural History and Science, in Albuqueque, New Mexico.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, Journal of Paleontology, Volume 35, 3rd issue, on pages 575-577.
- Gilmore, Charles W., On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, Band 38, Heft 43, Seiten 29-48, Parks. Veröffentlicht: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. und Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; und Dodson, Peter (eds.) The Dinosauria, 1st edition, Chapter: Hadrosauridae, Seiten 534-561. Veröffentlicht: 1990, University of California Press in Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., und Reisz, Robert R. 2007. Anatomie und Verwandtschaftsbeziehungen von Lambeosaurus magnicristatus, einem Hadrosauriden-Schopfsaurier (Ornithischia) aus der Dinosaur Park Formation, Alberta, aus dem Journal of Vertebrate Paleontology, Bd. 27 Ausgabe 2, auf den Seiten 373-393.
- Godefroit, Pascal, Shuqin Zan; und Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., ein lambeosauriner Dinosaurier aus dem späten Maastricht des nordöstlichen Chinas, aus den Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, Bd. 330, S. 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Published by New Mexico Museum of Natural History and Science, in Albuqueque, New Mexico. Seiten 191-213.
- Liddell, Henry George und Robert Scott, 1980. A Greek-English Lexicon (Gekürzte Ausgabe) Veröffentlicht: Oxford University Press in the United Kingdom. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. und Evans, David C., Currie, Phillip J., und Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, Kapitel: Ornithischian Dinosaurs. 2005, veröffentlicht: Indiana University Press, in Bloomington. Seiten 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; und Dupuis, Kevin, 2007. Ein juveniles Parasaurolophus-Gehirn aus dem Dinosaur Provincial Park, Alberta, mit Anmerkungen zur Ontogenese des Kammes in der Gattung. Journal of Vertebrate Paleontology, vol. 27, issue 3 pages 642-650.
- Currie, Phillip J; und Russell, Dale A. „Geographic and stratigraphic distribution of dinosaur remains“ in Dinosaur Provincial Park, S. 553.
- Weishampel, David B. (1990). „Dinosaurier-Verteilung“, in The Dinosauria (1.), S. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; und Noto, Christopher R. (2004). „Dinosaurier Distribution“, in The Dinosauria (2nd), pp. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, aus der Nova Acta Regia Societas Scientarum Upsaliensis, Serie 4, Bd. 7, Heft 5. (Deutsch). Seiten 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, veröffentlicht von Cassell Ltd. 1979, Ausgabe 5, in London. ISBN 0-304-52257-0 Seite 883.
- Weishampel, David B. und Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) aus Utah, aus dem Journal of Paleontology, Bd. 53, Heft 6, Seiten 1422-1427.
- Eberth, David A. 2005. „Die Geologie“, in Dinosaur Provincial Park, S. 54-82.
- Braman, Dennis R., und Koppelhus, Eva B. 2005. „Campanian palynomorphs“, in Dinosaur Provincial Park, pp. 101-130.
- Russell, Dale A. An Odyssey in Time: Dinosaurs of North America, 1989. Verlag: NorthWord Press, in Minocqua, Wisconsin. ISBN 1-55971-038-1 Seiten 160-164.
- Bakker, Robert T. 1986. The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, erschienen bei William Morrow, New York. ISBN 0-8217-2859-8 Seite 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: Ein origineller und fesselnder Einblick in das Leben im Dinosaurierreich, Kapitel: Hadrosauriden II. 1985. Veröffentlicht von Crescent Books in New York. Seiten 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs, aus dem Journal of Paleobiology, vol. 32, issue 1, Seiten 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs , aus dem Journal of Paleobiology, Band eins, Ausgabe eins, Seiten 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology, von der University of Chicago Press, auf Seite 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, aus der Zeitschrift Orion (München), Bd. 2. S. 525-532 (deutsch).
- Sternberg, Charles M. 1935. Hooded Hadrosaurs of the Belly River Series of the Upper Cretaceous aus dem Journal of the Canada Department of Mines Bulletin (Geological Series), Band 77, Heft 52, auf den Seiten 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, veröffentlicht 1945 vom American Museum of Natural History, Man and Nature Publications, 14, in New York. Seite 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas aus der Zeitschrift Jarbuch Naturwissenschaften, Band 12, Heft 36, auf den Seiten 709-716. (Deutsch) 1924.
- Osmólska, Halszka, 1979. Nasale Salzdrüsen bei Dinosauriern, aus der Zeitschrift Acta Palaeontologica Polonica, Bd. 24, S. 205-215.
- 36.0 36.1 Ostrom, John H., 1962. The cranial crests of hadrosaurian dinosaurs, aus: Journal of Postilla, Bd. 62, S. 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, veröffentlicht von Master Books, in Green Forest. ISBN 0-89051-165-9 Seite 82.
- 38.0 38.1 Weishampel, David B., 1981. Akustische Analysen der potentiellen Vokalisation bei lambeosaurinen Dinosauriern (Reptilia:Ornithischia), aus: Journal of Paleobiology, Bd. 7, Heft 2, S. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, aus dem Journal of Paleontology, vol. 55, issue 5, pp. 1046-1057.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur, von den Sandia National Laboratories (1997-12-05). Abgerufen am 20. Januar 2009.
- Diegert, Carl F. und Williamson, Thomas E., 1998. Ein digitales akustisches Modell des lambeosaurischen Hadrosauriers Parasaurolophus tubicen aus dem Journal of Vertebrate Paleontology, Bd. 18, Ausgabe 3, Suppl. S. 38A.
- Sullivan, Robert M. und Williamson, Thomas E., 1996. Ein neuer Schädel von Parasaurolophus (Langhaubenform) aus New Mexico: äußere und innere (CT-Scans) Merkmale und ihre funktionellen Auswirkungen, aus dem Journal of Vertebrate Paleontology, Bd. 16, Ausgabe 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborate CNS cooling structure in large dinosaurs Journal of Nature, vol. 275, on pp. 441-443.
- Maryańska, Teresa und Osmólska, Halszka, 1979. Aspekte der Schädelanatomie der Hadrosaurier, aus: Journal of Lethaia, Bd. 12, auf S. 265-273.
- Wissenschaftler verwenden digitale Paläontologie, um die Stimme des Dinosauriers Parasaurolophus zu erzeugen; von Sandia National Laboratories.
- Restaurationen von P. walkeri und P. cyrtocristatus, im gleichen Maßstab, von Scott Hartman; auf Skeletal Drawing.com.
- Parasaurolophus Seite auf Enchanted Learning.com