

Parasaurolophus (pronunciado /ˌpærəsɔˈrɒləfəs/ PARR-ə-sierra-ROL-ə-fəs, comúnmente también /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; que significa «lagarto cercano a la cresta» en referencia a Saurolophus) es un género de dinosaurio ornitópodo del período Cretácico Superior de lo que hoy es América del Norte, hace unos 76-73 millones de años. Era un herbívoro que caminaba tanto como bípedo como cuadrúpedo. Se reconocen tres especies: P. walkeri (la especie tipo), P. tubicen y el P. cyrtocristatus de cresta corta. Se conocen restos de Alberta, Canadá, y de Nuevo México y Utah, Estados Unidos. Fue descrito por primera vez en 1922 por William Parks a partir de un cráneo y un esqueleto parcial en Alberta.
Parasaurolophus es un hadrosáurido, parte de una diversa familia de dinosaurios del Cretácico conocidos por su gama de extraños adornos en la cabeza. Este género es conocido por su gran y elaborada cresta craneal, que en su mayor tamaño forma un largo tubo curvado que se proyecta hacia arriba y hacia atrás del cráneo. El Charonosaurus de China, que podría ser su pariente más cercano, tenía un cráneo similar y potencialmente una cresta similar. La cresta ha sido muy discutida por los científicos; el consenso es que las principales funciones incluían el reconocimiento visual tanto de la especie como del sexo, la resonancia acústica y la termorregulación. Es uno de los pico de pato más raros, ya que sólo se conoce a partir de un puñado de buenos especímenes.
Parasaurolophus se encuentra entre los dinosaurios más reconocidos, y se representa con frecuencia en programas de televisión relacionados con los dinosaurios, películas, libros, mercancías y otros medios de comunicación. Alice Perry
- Descripción
- Clasificación
- Historia
- Descubrimiento y denominación
- Especie
- Paleobiología
- Paleoecología
- Alimentación
- Cresta craneal
- Diferencias entre especies y etapas de crecimiento
- Hipótesis rechazadas sobre la función
- Funciones sociales
- Función sonora
- Función de refrigeración
- En los medios de comunicación
- Ver también
Descripción

El tamaño del Parasaurolophus walkeri comparado con el de un ser humano.
Como ocurre con la mayoría de los dinosaurios, el esqueleto del Parasaurolophus se conoce de forma incompleta. La longitud del espécimen tipo de P. walkeri se estima en 9,5 metros. Su cráneo mide aproximadamente 1,6 metros (5,2 pies) de largo, incluyendo la cresta, mientras que el cráneo tipo de P. tubicen mide más de 2,0 metros (6,6 pies) de largo, lo que indica un animal más grande. Su peso se estima en 2,5 toneladas. Su única extremidad delantera conocida era relativamente corta para un hadrosáurido, con un omóplato corto pero ancho. El hueso del muslo mide 103 centímetros (3,38 pies) de largo en P. walkeri y es robusto para su longitud en comparación con otros hadrosáuridos. La parte superior del brazo y los huesos de la pelvis también estaban fuertemente construidos.
Al igual que otros hadrosáuridos, era capaz de caminar en dos o cuatro patas. Probablemente prefería buscar comida en cuatro patas, pero corría en dos. Las espinas neurales de las vértebras eran altas, como era común en los lambeosaurinos; más altas sobre las caderas, aumentaban la altura de la espalda. Se conocen impresiones de la piel de P. walkeri, que muestran escamas uniformes parecidas a tubérculos, pero sin estructuras más grandes.
Parasaurolophus walkeri with scalation detail.
La característica más notable era la cresta craneal, que sobresalía de la parte posterior de la cabeza y estaba formada por el premaxilar y los huesos nasales. El espécimen tipo de P. walkeri tiene una muesca en las espinas neurales cerca de donde la cresta golpearía la espalda, pero puede tratarse de una patología propia de este individuo. William Parks, que dio nombre al género, planteó la hipótesis de que un ligamento iba desde la cresta hasta la muesca para sostener la cabeza. Aunque la idea parece poco probable, Parasaurolophus se restablece a veces con un colgajo de piel desde la cresta hasta el cuello.
La cresta era hueca, con tubos distintos que iban desde cada fosa nasal hasta el final de la cresta antes de invertir la dirección y volver a bajar por la cresta y entrar en el cráneo. Los tubos eran más simples en P. walkeri, y más complejos en P. tubicen, donde algunos tubos eran ciegos y otros se juntaban y separaban. Mientras que P. walkeri y P. tubicen tenían crestas largas con una ligera curvatura, P. cyrtocristatus tenía una cresta corta con un perfil más circular.
Clasificación
Como su nombre indica, inicialmente se pensó que Parasaurolophus estaba estrechamente relacionado con Saurolophus debido a su cresta superficialmente similar. Sin embargo, pronto se reevaluó como miembro de la subfamilia de los lambeosaurios, ya que Saurolophus es un hadrosaurio. Suele interpretarse como una rama separada de los lambeosaurinos, distinta de los Corythosaurus, Hypacrosaurus y Lambeosaurus con cresta. Su pariente más cercano conocido parece ser el Charonosaurus, un lambeosaurino con un cráneo similar (pero sin cresta completa todavía) de la región de Amur, en el noreste de China, y los dos pueden formar un clado Parasaurolophini. P. cyrtocristatus, con su cresta corta y más redondeada, puede ser la más basal de las tres especies conocidas de Parasaurolophus, o puede representar especímenes subadultos o hembras de P. tubicen.
Historia
Descubrimiento y denominación
Formación del espécimen tipo incompleto de Parasaurolophus walkeri en Varsovia.
Significa «lagarto con cresta cercana», el nombre Parasaurolophus deriva del griego para/παρα «al lado» o «cerca», saurus/σαυρος «lagarto» y lophos/λοφος «cresta». Se basa en el ROM 768, un cráneo y un esqueleto parcial al que le falta la mayor parte de la cola y las patas traseras por debajo de las rodillas, que fue encontrado por un grupo de campo de la Universidad de Toronto en 1920 cerca de Sand Creek a lo largo del río Red Deer en Alberta, Canadá. Estas rocas se conocen ahora como la Formación del Parque de los Dinosaurios del Cretácico Superior. William Parks bautizó el espécimen como P. walkeri en honor a Sir Byron Edmund Walker, presidente del Consejo de Administración del Museo Real de Ontario. Los restos de Parasaurolophus son raros en Alberta, con sólo otro cráneo parcial de (probablemente) la Formación Dinosaur Park, y tres especímenes de Dinosaur Park que carecen de cráneo, posiblemente pertenecientes al género. En algunas listas de fauna, se menciona un posible material de P. walkeri en la Formación Hell Creek de Montana, una unidad de roca de edad maastrichtiana tardía. Sullivan y Williamson no mencionan esta presencia en su revisión del género de 1999, y no se ha profundizado en ella en otros lugares.
En 1921, Charles H. Sternberg recuperó un cráneo parcial (PMU.R1250) de lo que ahora se conoce como la Formación Kirtland, ligeramente más joven, en el condado de San Juan, Nuevo México. Este espécimen fue enviado a Uppsala, Suecia, donde Carl Wiman lo describió como una segunda especie, P. tubicen, en 1931. El epíteto específico deriva del latín tǔbǐcěn «trompetista». Un segundo cráneo casi completo de P. tubicen (NMMNH P-25100) fue encontrado en Nuevo México en 1995. Utilizando la tomografía computarizada de este cráneo, Robert Sullivan y Thomas Williamson dieron al género un tratamiento monográfico en 1999, cubriendo aspectos de su anatomía y taxonomía, y las funciones de su cresta. Williamson publicó posteriormente una revisión independiente de los restos, discrepando de las conclusiones taxonómicas.
John Ostrom describió otro buen ejemplar (FMNH P27393) de Nuevo México como P. cyrtocristatus en 1961. Incluye un cráneo parcial con una cresta corta y redondeada, y gran parte del esqueleto postcraneal, excepto los pies, el cuello y partes de la cola. Su nombre específico deriva del latín curtus «acortado» y cristatus «crestado». El espécimen se encontró en la parte superior de la formación Fruitland o, más probablemente, en la base de la formación Kirtland. El área de distribución de esta especie se amplió en 1979, cuando David B. Weishampel y James A. Jensen describieron un cráneo parcial con una cresta similar (BYU 2467) de la Formación Kaiparowits, de edad campaniana, del condado de Garfield, Utah. Desde entonces, se ha encontrado otro cráneo en Utah con la morfología de cresta corta/redonda de P. cyrtocristatus.
Especie
El molde del cráneo de Parasaurolophus walkeri, Museo de Historia Natural, Londres.
Modelo de cráneo de Parasaurolophus cyrtocristatus, Field Museum of Natural History.
La especie tipo P. walkeri, de Alberta, se conoce a partir de un único ejemplar. Se diferencia de P. tubicen por tener tubos más simples en su cresta, y de P. cyrtocristatus por tener una cresta larga y no redondeada y un brazo superior más largo que el antebrazo.
P. tubicen, de Nuevo México, se conoce por los restos de al menos tres individuos. Es la especie más grande, con pasajes de aire más complejos en su cresta que P. walkeri, y una cresta más larga y recta que P. cyrtocristatus.
P. cyrtocristatus, de Nuevo México y Utah, se conoce a partir de tres posibles especímenes. Es la especie más pequeña, con una cresta corta y redondeada. Su pequeño tamaño y la forma de su cresta han llevado a varios científicos a sugerir que representa a juveniles o hembras de P. tubicen, que es de aproximadamente la misma época y de la misma formación en Nuevo México. Como señaló Thomas Williamson, el material tipo de P. cyrtocristatus tiene aproximadamente un 72% del tamaño de P. tubicen, cerca del tamaño en el que se interpreta que otros lambeosaurinos comienzan a mostrar un dimorfismo sexual definitivo en sus crestas (~70% del tamaño adulto). Esta posición ha sido rechazada en recientes revisiones de lambeosaurinos.
Paleobiología
Paleoecología
Gorgosaurus (izquierda) atacando a un Parasaurolophus cyrtocristatus.
Parasaurolophus walkeri, de la Formación Dinosaur Park, fue miembro de una diversa y bien documentada fauna de animales prehistóricos, entre los que se encuentran conocidos dinosaurios como el Centrosaurus con cuernos, el Styracosaurus y el Chasmosaurus; sus compañeros Prosaurolophus, Gryposaurus, Corythosaurus y Lambeosaurus; el tiranosáurido Gorgosaurus; y los acorazados Edmontonia y Euoplocephalus. Era un constituyente poco frecuente de esta fauna. La Formación Parque de los Dinosaurios se interpreta como un entorno de bajo relieve de ríos y llanuras aluviales que con el tiempo se volvió más pantanoso e influenciado por las condiciones marinas a medida que la vía marítima interior occidental transgredía hacia el oeste. El clima era más cálido que el de la actual Alberta, sin heladas, pero con estaciones más húmedas y más secas. Las coníferas eran aparentemente las plantas dominantes del dosel, con un sotobosque de helechos, helechos arborescentes y angiospermas.
Las especies de Nuevo México compartieron su entorno con el gran saurópodo Alamosaurus, el Kritosaurus con pico de pato, el Pentaceratops con cuernos, el Nodocephalosaurus con armadura, el Saurornitholestes y tiranosáuridos actualmente sin nombre. La Formación Kirtland se interpreta como llanuras de inundación fluvial que aparecieron tras el retroceso de la vía marítima interior occidental. Las coníferas eran las plantas dominantes, y los dinosaurios con cuernos chasmosaurinos eran aparentemente más comunes que los hadrosáuridos.
Alimentación
Como hadrosáurido, Parasaurolophus era un gran herbívoro bípedo/cuadrúpedo, que comía plantas con un sofisticado cráneo que permitía un movimiento de trituración análogo al de la masticación. Sus dientes se reemplazaban continuamente y se empaquetaban en baterías dentales que contenían cientos de dientes, de los cuales sólo un relativo puñado estaba en uso en cualquier momento. Utilizaba el pico para cosechar material vegetal, que era sujetado en las mandíbulas por un órgano parecido a una mejilla. Se alimentaba desde el suelo hasta unos 4 metros de altura. Como señaló Bob Bakker, los lambeosaurinos tienen picos más estrechos que los hadrosaurinos, lo que implica que Parasaurolophus y sus parientes podían alimentarse de forma más selectiva que sus homólogos de pico ancho y sin cresta.
Cresta craneal
Se han propuesto muchas hipótesis sobre las funciones que desempeñaba la cresta craneal de Parasaurolophus, pero la mayoría han sido desacreditadas. En la actualidad se cree que pudo tener varias funciones: visualización para identificar la especie y el sexo, amplificación del sonido para la comunicación y termorregulación. No está claro cuál fue la más importante y en qué momento de la evolución de la cresta y sus conductos nasales internos.
Diferencias entre especies y etapas de crecimiento
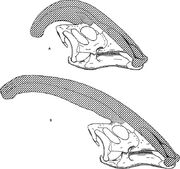
Diagrama comparativo de las crestas nariales de Parasaurolophus cyrtocristatus (a) y Parasaurolophus walkeri (b).
Al igual que otros lambeosaurinos, se cree que la cresta craneal de Parasaurolophus cambiaba con la edad y era una característica sexualmente dimórfica en los adultos. James Hopson, uno de los primeros investigadores en describir las crestas de los lambeosaurinos en términos de tales distinciones, sugirió que P. cyrtocristatus, con su pequeña cresta, era la forma femenina de P. tubicen. Thomas Williamson sugirió que era la forma juvenil. Ninguna de las dos hipótesis fue ampliamente aceptada. Como sólo se conocen seis cráneos en buen estado y una caja de cerebros juvenil, el material adicional ayudará a aclarar estas posibles relaciones. Williamson señaló que, en cualquier caso, los Parasaurolophus juveniles probablemente tenían crestas pequeñas y redondeadas, como P. cyrtocristatus, que probablemente crecían más rápido a medida que los individuos se acercaban a la madurez. El reciente reestudio de una caja de cerebros juvenil previamente asignada a Lambeosaurus, y ahora asignada a Parasaurolophus, proporciona pruebas de que una pequeña cresta tubular estaba presente en los juveniles. Este espécimen conserva un pequeño ensanchamiento hacia arriba de los huesos frontales que era similar pero más pequeño que lo que se ve en los especímenes adultos; en los adultos, los frontales formaban una plataforma que sostenía la base de la cresta. Este espécimen también indica que el crecimiento de la cresta en Parasaurolophus y el perfil facial de los individuos juveniles difería del modelo Corythosaurus-Hypacrosaurus-Lambeosaurus, en parte porque la cresta de Parasaurolophus carece del delgado «coxcomb» óseo que constituye la parte superior de la cresta de los otros tres lambeosaurinos.
Hipótesis rechazadas sobre la función
Muchas de las primeras sugerencias se centraron en las adaptaciones para un estilo de vida acuático, siguiendo la hipótesis de que los hadrosáuridos eran anfibios, una línea de pensamiento común hasta la década de 1960. Así, Alfred Sherwood Romer propuso que servía como esnórquel, Martin Wilfarth que era un accesorio para una probóscide móvil utilizada como tubo de respiración o para la recolección de alimentos, Charles M. Sternberg que servía como trampa de aire para mantener el agua fuera de los pulmones, y Ned Colbert que servía como depósito de aire para estancias prolongadas bajo el agua.
Otras propuestas eran de naturaleza más física. Como se ha mencionado anteriormente, William Parks sugirió que estaba unida a las vértebras con ligamentos o músculos, y ayudaba a mover y sostener la cabeza. Othenio Abel propuso que se utilizaba como arma en los combates entre miembros de la misma especie, y Andrew Milner sugirió que podía utilizarse como deflector de follaje, como la cresta del casco (llamada «casque») del casuario. Otras propuestas hacían que la función principal fuera la de albergar órganos especializados. Halszka Osmólska sugirió que albergaba glándulas de sal, y John Ostrom sugirió que albergaba zonas ampliadas para el tejido olfativo y un sentido del olfato muy mejorado de los lambeosaurinos, que no tenían capacidades defensivas evidentes. Una sugerencia inusual, hecha por el creacionista Duane Gish, es que la cresta albergaba glándulas químicas que le permitían lanzar chorros de «fuego» químico a los enemigos, similar al escarabajo bombardero actual.
La mayoría de estas hipótesis han sido desacreditadas o rechazadas. Por ejemplo, no hay ningún agujero en el extremo de la cresta para una función de esnórquel. No hay cicatrices musculares para una probóscide y es dudoso que un animal con pico la necesite. Como esclusa propuesta, no habría impedido la entrada de agua. El depósito de aire propuesto habría sido insuficiente para un animal del tamaño de Parasaurolophus. Otros hadrosáuridos tenían cabezas grandes sin necesidad de grandes crestas huecas que sirvieran de puntos de unión para los ligamentos de soporte. Además, ninguna de las propuestas explica por qué la cresta tiene esa forma, por qué otros lambeosaurinos deberían tener crestas de aspecto muy diferente pero que cumplen una función similar, cómo los hadrosáuridos sin cresta o con cresta sólida se las arreglaban sin esas capacidades, o por qué algunos hadrosáuridos tenían crestas sólidas. Estas consideraciones afectan especialmente a las hipótesis basadas en el aumento de las capacidades de sistemas ya presentes en el animal, como las hipótesis de la glándula de la sal y la olfacción, e indican que éstas no eran funciones primarias de la cresta. Además, los trabajos sobre la cavidad nasal de los lambeosaurinos muestran que los nervios olfativos y el tejido sensorial correspondiente estaban en gran medida fuera de la porción de las fosas nasales de la cresta, por lo que la expansión de la cresta tenía poco que ver con el sentido del olfato.
Restauración de la vida de Parasaurolophus walkeri con un hipotético volante de piel y rayas en su cresta.
En cambio, las funciones sociales y fisiológicas se han apoyado más como función(es) de la cresta, centrándose en la identificación y comunicación visual y auditiva. Como objeto de gran tamaño, la cresta tiene un claro valor como señal visual, y diferencia a este animal de sus contemporáneos. El gran tamaño de las cuencas oculares de los hadrosáuridos y la presencia de anillos escleróticos en los ojos implican una visión aguda y hábitos diurnos, prueba de que la vista era importante para estos animales. Si, como se suele ilustrar, un volante de piel se extendía desde la cresta hasta el cuello o la espalda, el despliegue visual propuesto habría sido mucho más vistoso. Como sugieren otros cráneos de lambeosaurinos, la cresta de Parasaurolophus probablemente permitía tanto la identificación de la especie (como separarla de Corythosaurus o Lambeosaurus) como la determinación entre machos y hembras, basándose en la forma y el tamaño.
Función sonora
Modelo informático de los conductos nasales de la cresta de P. tubicen.
Sin embargo, el aspecto externo de la cresta no se corresponde con la compleja anatomía interna de los conductos nasales, lo que sugiere otra función que explica el uso del espacio interno. Carl Wiman fue el primero en proponer, en 1931, que los conductos cumplían una función de señalización auditiva, como un claxon; Hopson y David B. Weishampel retomaron esta idea en las décadas de 1970 y 1980. Hopson descubrió que existen pruebas anatómicas de que los hadrosáuridos tenían una gran capacidad auditiva. Hay al menos un ejemplo, en el afín Corythosaurus, de un estribo delgado (hueso del oído de los reptiles) en su lugar, que combinado con un gran espacio para un tímpano implica un oído medio sensible. Además, la lagena de los hadrosaurios es alargada como la de los cocodrilos, lo que indica que la parte auditiva del oído interno estaba bien desarrollada. Weishampel sugirió que P. walkeri era capaz de producir frecuencias de 48 a 240 Hz, y P. cyrtocristatus (interpretado como una forma juvenil de cresta) de 75 a 375 Hz. Basándose en la similitud de los oídos internos de los hadrosáuridos con los de los cocodrilos, también propuso que los hadrosáuridos adultos eran sensibles a las altas frecuencias, como las que podrían producir sus crías. Según Weishampel, esto es coherente con la comunicación entre padres y crías.
El modelado por ordenador de un espécimen bien conservado de P. tubicen, con conductos de aire más complejos que los de P. walkeri, ha permitido reconstruir el posible sonido que producía su cresta. El trayecto principal resuena a unos 30 Hz, pero la complicada anatomía del seno provoca picos y valles en el sonido.
Función de refrigeración
La gran superficie y la vascularización de la cresta también sugieren una función termorreguladora. P.E. Wheeler sugirió por primera vez este uso en 1978 como forma de mantener fresco el cerebro. Teresa Maryańska y Osmólska también propusieron la termorregulación casi al mismo tiempo, y Sullivan y Williamson se interesaron aún más. La discusión de David Evans en 2006 sobre las funciones de la cresta de los lambeosaurios fue favorable a la idea, al menos como factor inicial para la evolución de la expansión de la cresta.
En los medios de comunicación
El Parasaurolophus es un dinosaurio muy popular. Apareció en la película Dinosaurio de Disney como miembro de la manada. También hizo varias apariciones en el famoso documental Clash of the Dinosaurs. También apareció en el popular programa de televisión Prehistoric Park, donde se convirtió en la presa de los grandes carnívoros Deinosuchus y Albertosaurus. Hizo algunas apariciones en las películas de Parque Jurásico, como miembro de la manada en la primera película, siendo cautivo de los cazadores en la segunda y corriendo junto al Corythosaurus en la tercera. y el personaje Dweeb en «¡Hemos vuelto! A Dinosaur’s Story», es el propio Parasaurolophus.
El Parasaurolophus también aparece en Turok, como un dócil comedor de plantas que normalmente no es dañino, pero que puede ser agresivo si se le provoca gravemente.
También hay un zord Parasaurolophus en Power Rangers: Dino Thunder y Power Rangers: Dino Charge.
El Parasaurolophus aparece brevemente al principio de la película de Disney Pixar The Good Dinosaur.
Ver también
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, página 229. Publicado: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Chapter: Parasaurolophus, páginas 678-684. Publicado: 1997, McFarland & Co, en Jefferson, Carolina del Norte. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull y Wright, Hadrosaurian Dinosaurs of North America, pp. 209-213.
- Brett-Surman, Michael K. y Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, capítulo: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, páginas 135-169. Publicado, 2006, Indiana University Press, en Bloomington e Indianápolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; y Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; y Dodson, Peter (eds.) The Dinosauria, 2ª edición, capítulo: Hadrosauridae, páginas 438-463. Publicado: 2004, University of California Press, en Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, volume 13, pages 1-32. Publicado: 1922, Estudios de la Universidad de Toronto, Serie de Geología.
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. and Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, from the series New Mexico Museum of Natural History and Science Bulletin, 15, pages 1-52. Publicado: 1999, Museo de Historia Natural y Ciencia de Nuevo México, en Albuqueque, Nuevo México.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, Journal of Paleontology, Volume 35, 3rd issue, on pages 575-577.
- Gilmore, Charles W., Sobre el género Stephanosaurus, con una descripción del espécimen tipo de Lambeosaurus lambei, volumen 38, número 43, páginas 29-48, Parks. Publicado: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. y Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; y Dodson, Peter (eds.) The Dinosauria, 1st edition, Chapter: Hadrosauridae, páginas 534-561. Publicado: 1990, Universidad de California Press en Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., y Reisz, Robert R. 2007. Anatomía y relaciones de Lambeosaurus magnicristatus, un dinosaurio hadrosáurido con cresta (Ornithischia) de la Formación del Parque de los Dinosaurios, Alberta, del Journal of Vertebrate Paleontology, vol. 27 número 2, en las páginas 373-393.
- Godefroit, Pascal, Shuqin Zan; y Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China, from the Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, pages 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Revisión de Hadrosauridae (Dinosauria, Ornithischia) de la Cuenca de San Juan, Nuevo México Lucas, S.G.; y Heckert, A.B. (eds.) Dinosaurios de Nuevo México, del Museo de Nuevo México de Historia Natural y Ciencia Boletín, 17 Publicado por el Museo de Nuevo México de Historia Natural y Ciencia, en Albuqueque, Nuevo México. Páginas 191-213.
- Liddell, Henry George y Robert Scott, 1980. A Greek-English Lexicon (Edición abreviada) Publicado: Oxford University Press en el Reino Unido. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. y Evans, David C., Currie, Phillip J., y Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, Chapter: Ornithischian Dinosaurs. 2005, publicado: Indiana University Press, en Bloomington. Páginas 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; y Dupuis, Kevin, 2007. A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus. Journal of Vertebrate Paleontology, vol. 27, número 3 páginas 642-650.
- Currie, Phillip J; y Russell, Dale A. «Geographic and stratigraphic distribution of dinosaur remains» in Dinosaur Provincial Park, p. 553.
- Weishampel, David B. (1990). «Distribución de los dinosaurios», en The Dinosauria (1), pp. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; y Noto, Christopher R. (2004). «Dinosaur Distribution», en The Dinosauria (2nd), pp. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, de la Nova Acta Regia Societas Scientarum Upsaliensis, serie 4, vol. 7, número 5. (alemán). Páginas 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, publicado por Cassell Ltd. 1979, edición 5, en Londres. ISBN 0-304-52257-0 Página 883.
- Weishampel, David B. y Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) de Utah, del Journal of Paleontology, vol. 53, número 6, páginas 1422-1427.
- Eberth, David A. 2005. «La geología», en el Parque Provincial de los Dinosaurios, pp. 54-82.
- Braman, Dennis R., y Koppelhus, Eva B. 2005. «Campanian palynomorphs», en Dinosaur Provincial Park, pp. 101-130.
- Russell, Dale A. An Odyssey in Time: Dinosaurios de América del Norte, 1989. Editorial: NorthWord Press, en Minocqua, Wisconsin. ISBN 1-55971-038-1 Páginas 160-164.
- Bakker, Robert T. 1986. Las herejías de los dinosaurios: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, publicado por William Morrow, en Nueva York. ISBN 0-8217-2859-8 Página 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom, capítulo: Hadrosaurios II. 1985. Publicado por Crescent Books, en Nueva York. Páginas 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Homologías de la cavidad nasal y la función de la cresta craneal en los dinosaurios lambeosaurinos, del Journal of Paleobiology, vol. 32, número 1, Páginas 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. La evolución de las estructuras de visualización craneal en los dinosaurios hadrosaurios , de la Revista de Paleobiología, volumen uno, número uno, páginas 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology, de la University of Chicago Press, en la página 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, del Journal of Orion (Munich), vol. 2. pp. 525-532 (en alemán).
- Sternberg, Charles M. 1935. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous from the Journal of the Canada Department of Mines Bulletin (Geological Series), volume 77, issue 52, on pages 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, publicado en 1945 por el Museo Americano de Historia Natural, Man and Nature Publications, 14, en Nueva York. Página 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas del Journal of Jarbuch Naturwissenschaften, volumen 12, número 36, en las páginas 709-716. (Alemán) 1924.
- Osmólska, Halszka, 1979. Glándulas nasales de sal en los dinosaurios, de la Revista de Acta Palaeontologica Polonica, vol. 24, páginas 205-215.
- 36.0 36.1 Ostrom, John H., 1962. Las crestas craneales de los dinosaurios hadrosaurios, del Journal of Postilla, vol. 62, páginas 1-29.
- Gish, Duane T., 1992. Dinosaurios por diseño, publicado por Master Books, en Green Forest. ISBN 0-89051-165-9 Página 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), del Journal of Paleobiology, vol. 7, número 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, from the Journal of Paleontology, vol. 55, issue 5, pp. 1046-1057.
- Los científicos utilizan la paleontología digital para producir la voz del dinosaurio Parasaurolophus, de los Sandia National Laboratories (1997-12-05). Recuperado el 20 de enero de 2009.
- Diegert, Carl F. y Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen from the Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. y Williamson, Thomas E., 1996. A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications, from the Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborada estructura de enfriamiento del SNC en los grandes dinosaurios Journal of Nature, vol. 275, en pp. 441-443.
- Maryańska, Teresa y Osmólska, Halszka, en 1979. Aspectos de la anatomía craneal de los hadrosaurios, de la Revista de Lethaia, vol. 12, en las páginas 265-273.
- Los científicos utilizan la paleontología digital para producir la voz del dinosaurio Parasaurolophus; de Sandia National Laboratories.
- Restauraciones de P. walkeri y P. cyrtocristatus, a la misma escala, por Scott Hartman; en Skeletal Drawing.com.
- Página de Parasaurolophus en Enchanted Learning.com