

Parasaurolophus (prononcé /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, communément aussi /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs ; signifiant » lézard à crête » en référence à Saurolophus) est un genre de dinosaure ornithopode du Crétacé supérieur de ce qui est aujourd’hui l’Amérique du Nord, il y a environ 76 à 73 millions d’années. C’était un herbivore qui marchait à la fois comme un bipède et un quadrupède. Trois espèces sont reconnues : P. walkeri (l’espèce type), P. tubicen, et le P. cyrtocristatus à crête courte. Des vestiges sont connus en Alberta, au Canada, ainsi qu’au Nouveau-Mexique et en Utah, aux États-Unis. Il a été décrit pour la première fois en 1922 par William Parks à partir d’un crâne et d’un squelette partiel en Alberta.
Parasaurolophus est un hadrosauridé, faisant partie d’une famille diverse de dinosaures du Crétacé connus pour leur gamme d’ornements de tête bizarres. Ce genre est connu pour sa grande crête crânienne élaborée, qui, dans sa plus grande taille, forme un long tube incurvé faisant saillie vers le haut et l’arrière du crâne. Le Charonosaurus de Chine, qui pourrait être son plus proche parent, avait un crâne similaire et potentiellement une crête similaire. La crête a fait l’objet de nombreuses discussions entre les scientifiques ; le consensus est que ses principales fonctions comprenaient la reconnaissance visuelle de l’espèce et du sexe, la résonance acoustique et la thermorégulation. C’est l’un des becs de canard les plus rares, n’étant connu que par une poignée de bons spécimens.
Parasaurolophus est parmi les dinosaures les plus reconnus, et est fréquemment représenté dans les émissions de télévision, les films, les livres, les marchandises et autres médias liés aux dinosaures. Alice Perry
Description

La taille de Parasaurolophus walkeri par rapport à un humain.
Comme c’est le cas pour la plupart des dinosaures, le squelette de Parasaurolophus est incomplètement connu. La longueur du spécimen type de P. walkeri est estimée à 9,5 mètres (31 ft). Son crâne mesure environ 1,6 mètres (5,2 ft) de long, y compris la crête, alors que le crâne type de P. tubicen mesure plus de 2,0 mètres (6,6 ft) de long, ce qui indique un animal plus grand. Son poids est estimé à 2,5 tonnes (2,7 tonnes). Son unique membre antérieur connu était relativement court pour un hadrosauridé, avec une omoplate courte mais large. Le fémur mesure 103 centimètres (3,38 pieds) de long chez P. walkeri et est robuste pour sa longueur par rapport aux autres hadrosauridés. La partie supérieure du bras et les os du bassin étaient également fortement construits.
Comme les autres hadrosauridés, il était capable de marcher sur deux ou quatre pattes. Il préférait probablement chercher sa nourriture sur quatre pattes, mais courait sur deux. Les épines neurales des vertèbres étaient hautes, comme c’était courant chez les lambeosaurinés ; plus hautes au-dessus des hanches, elles augmentaient la hauteur du dos. Des empreintes de peau sont connues pour P. walkeri, montrant des écailles uniformes en forme de tubercules, mais pas de structures plus grandes.
Parasaurolophus walkeri avec détail d’écaillage.
La caractéristique la plus remarquable était la crête crânienne, qui dépassait de l’arrière de la tête et était composée du prémaxillaire et des os nasaux. Le spécimen type de P. walkeri présente une encoche dans les épines neurales près de l’endroit où la crête toucherait le dos, mais il peut s’agir d’une pathologie propre à cet individu. William Parks, qui a nommé le genre, a émis l’hypothèse qu’un ligament allait de la crête à l’encoche pour soutenir la tête. Bien que l’idée semble peu probable, Parasaurolophus est parfois restitué avec un lambeau de peau de la crête au cou.
La crête était creuse, avec des tubes distincts menant de chaque narine à l’extrémité de la crête avant d’inverser la direction et de se diriger à nouveau vers le bas de la crête et dans le crâne. Les tubes étaient les plus simples chez P. walkeri, et plus complexes chez P. tubicen, où certains tubes étaient aveugles et d’autres se rencontraient et se séparaient. Alors que P. walkeri et P. tubicen avaient de longues crêtes avec seulement une légère courbure, P. cyrtocristatus avait une crête courte avec un profil plus circulaire.
Classification
Comme son nom l’indique, Parasaurolophus était initialement considéré comme étroitement lié à Saurolophus en raison de sa crête superficiellement similaire. Cependant, il a rapidement été réévalué comme un membre de la sous-famille lambeosaurine des hadrosauridés-Saurolophus est un hadrosauridé. On l’interprète généralement comme une ramification distincte des lambeosaurins, différente du Corythosaurus, de l’Hypacrosaurus et du Lambeosaurus à crête. Son plus proche parent connu semble être Charonosaurus, un lambeosaurin avec un crâne similaire (mais pas encore de crête complète) de la région d’Amur au nord-est de la Chine, et les deux pourraient former un clade Parasaurolophini. P. cyrtocristatus, avec sa crête courte et plus ronde, pourrait être la plus basale des trois espèces connues de Parasaurolophus, ou représenter des spécimens subadultes ou femelles de P. tubicen.
Histoire
Découverte et dénomination
Coulée du spécimen type incomplet de Parasaurolophus walkeri à Warszawa.
Signifiant « lézard à crête proche », le nom Parasaurolophus est dérivé du grec para/παρα « à côté » ou « proche », saurus/σαυρος « lézard » et lophos/λοφος « crête ». Il est basé sur le ROM 768, un crâne et un squelette partiel auquel il manque la majeure partie de la queue et les pattes arrière sous les genoux, qui a été trouvé par une équipe de terrain de l’Université de Toronto en 1920 près de Sand Creek le long de la rivière Red Deer en Alberta, au Canada. Ces roches sont maintenant connues sous le nom de Formation Dinosaur Park du Crétacé supérieur, datant du Campanien. William Parks a nommé le spécimen P. walkeri en l’honneur de Sir Byron Edmund Walker, président du conseil d’administration du Royal Ontario Museum. Les restes de Parasaurolophus sont rares en Alberta, avec seulement un autre crâne partiel provenant (probablement) de la Formation de Dinosaur Park, et trois spécimens de Dinosaur Park dépourvus de crâne, appartenant peut-être au genre. Dans certaines listes de faune, il y a une mention de matériel possible de P. walkeri dans la Formation Hell Creek du Montana, une unité de roche d’âge Maastrichtien tardif. Cette occurrence n’est pas notée par Sullivan et Williamson dans leur revue du genre de 1999, et n’a pas été élaborée ailleurs.
En 1921, Charles H. Sternberg a récupéré un crâne partiel (PMU.R1250) de ce qui est maintenant connu comme la formation légèrement plus jeune de Kirtland dans le comté de San Juan, au Nouveau-Mexique. Ce spécimen a été envoyé à Uppsala, en Suède, où Carl Wiman l’a décrit comme une deuxième espèce, P. tubicen, en 1931. L’épithète spécifique est dérivée du latin tǔbǐcěn « trompette ». Un deuxième crâne de P. tubicen presque complet (NMMNH P-25100) a été trouvé au Nouveau-Mexique en 1995. En utilisant la tomographie assistée par ordinateur de ce crâne, Robert Sullivan et Thomas Williamson ont donné au genre un traitement monographique en 1999, couvrant les aspects de son anatomie et de sa taxonomie, ainsi que les fonctions de sa crête. Williamson a ensuite publié une revue indépendante des restes, en désaccord avec les conclusions taxonomiques.
John Ostrom a décrit un autre bon spécimen (FMNH P27393) du Nouveau-Mexique comme P. cyrtocristatus en 1961. Il comprend un crâne partiel avec une crête courte et arrondie, et une grande partie du squelette postcrânien à l’exception des pieds, du cou et de certaines parties de la queue. Son nom spécifique est dérivé du latin curtus « raccourci » et cristatus « crête ». Le spécimen a été trouvé soit dans le sommet de la Formation de Fruitland, soit, plus probablement, dans la base de la Formation de Kirtland sus-jacente. La gamme de cette espèce a été étendue en 1979, quand David B. Weishampel et James A. Jensen ont décrit un crâne partiel avec une crête semblable (BYU 2467) de la formation de Kaiparowits de l’âge Campanien de Garfield County, Utah. Depuis, un autre crâne a été trouvé dans l’Utah avec la morphologie de crête courte/ronde de P. cyrtocristatus.
Espèce
Moulage de crâne de Parasaurolophus walkeri, Musée d’histoire naturelle, Londres.
Parasaurolophus cyrtocristatus moulage de crâne, Field Museum of Natural History.
L’espèce type P. walkeri, de l’Alberta, est connue à partir d’un seul spécimen. Elle diffère de P. tubicen par des tubes plus simples dans sa crête, et de P. cyrtocristatus par une crête longue et non arrondie et un bras supérieur plus long que l’avant-bras.
P. tubicen, du Nouveau-Mexique, est connue par les restes d’au moins trois individus. C’est la plus grande espèce, avec des passages d’air plus complexes dans sa crête que P. walkeri, et une crête plus longue et plus droite que P. cyrtocristatus.
P. cyrtocristatus, du Nouveau-Mexique et de l’Utah, est connu à partir de trois spécimens possibles. C’est la plus petite espèce, avec une crête courte et arrondie. Sa petite taille et la forme de sa crête ont conduit plusieurs scientifiques à suggérer qu’elle représente des juvéniles ou des femelles de P. tubicen, qui date à peu près de la même époque et de la même formation au Nouveau-Mexique. Comme l’a noté Thomas Williamson, le matériel type de P. cyrtocristatus est environ 72% de la taille de P. tubicen, proche de la taille à laquelle d’autres lambeosaurines sont interprétées comme commençant à montrer un dimorphisme sexuel définitif dans leurs crêtes (~70% de la taille adulte). Cette position a été rejetée dans des revues récentes de lambeosaurines.
Paléobiologie
Paléoécologie
Gorgosaurus (à gauche) attaquant un Parasaurolophus cyrtocristatus.
Parasaurolophus walkeri, de la formation de Dinosaur Park, était un membre d’une faune diverse et bien documentée d’animaux préhistoriques, y compris des dinosaures bien connus comme le Centrosaurus cornu, le Styracosaurus et le Chasmosaurus ; les autres becs de canard Prosaurolophus, Gryposaurus, Corythosaurus et Lambeosaurus ; le tyrannosaure Gorgosaurus ; et les blindés Edmontonia et Euoplocephalus. C’était un constituant rare de cette faune. La formation de Dinosaur Park est interprétée comme un cadre de bas-relief de rivières et de plaines d’inondation qui sont devenues plus marécageuses et influencées par des conditions marines au fil du temps alors que la voie maritime intérieure occidentale transgressait vers l’ouest. Le climat était plus chaud que celui de l’Alberta actuelle, sans gelée, mais avec des saisons plus humides et plus sèches. Les conifères étaient apparemment les plantes dominantes de la canopée, avec un sous-étage de fougères, de fougères arborescentes et d’angiospermes.
Les espèces du Nouveau-Mexique partageaient leur environnement avec le grand sauropode Alamosaurus, le Kritosaurus à bec de canard, le Pentaceratops cornu, le Nodocephalosaurus blindé, le Saurornitholestes et des tyrannosauridés actuellement sans nom. La formation de Kirtland est interprétée comme des plaines d’inondation fluviales apparues après le retrait de la voie maritime intérieure occidentale. Les conifères étaient les plantes dominantes, et les dinosaures à cornes chasmosaurins étaient apparemment plus communs que les hadrosauridés.
Alimentation
En tant qu’hadrosauridé, Parasaurolophus était un grand herbivore bipède/quadrupède, mangeant des plantes avec un crâne sophistiqué qui permettait un mouvement de broyage analogue à la mastication. Ses dents étaient continuellement remplacées et emballées dans des batteries dentaires qui contenaient des centaines de dents, dont seule une poignée relative était utilisée à tout moment. Il utilisait son bec pour récolter les matières végétales, qui étaient maintenues dans les mâchoires par un organe ressemblant à une joue. Il se nourrissait depuis le sol jusqu’à environ 4 mètres (13 pieds) de hauteur. Comme l’a noté Bob Bakker, les lambeosaurinés ont des becs plus étroits que les hadrosaurinés, ce qui implique que Parasaurolophus et ses proches pouvaient se nourrir de manière plus sélective que leurs homologues à large bec et sans crête.
Crête crânienne
De nombreuses hypothèses ont été avancées quant aux fonctions que remplissait la crête crânienne de Parasaurolophus, mais la plupart ont été discréditées. On pense maintenant qu’elle pouvait avoir plusieurs fonctions : affichage visuel pour identifier l’espèce et le sexe, amplification du son pour la communication, et thermorégulation. On ne sait pas clairement laquelle était la plus importante à quels moments de l’évolution de la crête et de ses passages nasaux internes.
Différences entre espèces et stades de croissance
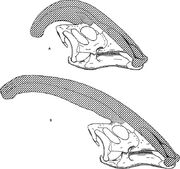
Diagramme comparant les crêtes nariales de Parasaurolophus cyrtocristatus (a) et de Parasaurolophus walkeri (b).
Comme pour les autres lambeosaurines, on pense que la crête crânienne de Parasaurolophus changeait avec l’âge et était une caractéristique sexuellement dimorphique chez les adultes. James Hopson, l’un des premiers chercheurs à décrire les crêtes des lambéosaurins en termes de telles distinctions, a suggéré que P. cyrtocristatus, avec sa petite crête, était la forme féminine de P. tubicen. Thomas Williamson a suggéré qu’il s’agissait de la forme juvénile. Aucune des deux hypothèses n’a été largement acceptée. Comme on ne connaît que six bons crânes et une boîte crânienne juvénile, du matériel supplémentaire permettra de clarifier ces relations potentielles. Williamson a noté que dans tous les cas, les Parasaurolophus juvéniles avaient probablement des crêtes petites et arrondies comme P. cyrtocristatus, qui se développaient probablement plus rapidement à mesure que les individus approchaient de la maturité. La réétude récente d’une boîte crânienne juvénile précédemment attribuée à Lambeosaurus, et maintenant attribuée à Parasaurolophus, fournit la preuve qu’une petite crête tubulaire était présente chez les juvéniles. Ce spécimen préserve un petit évasement vers le haut des os frontaux qui était similaire mais plus petit que ce qui est observé dans les spécimens adultes ; chez les adultes, les os frontaux formaient une plate-forme qui soutenait la base de la crête. Ce spécimen indique également que la croissance de la crête chez Parasaurolophus et le profil facial des individus juvéniles différaient du modèle Corythosaurus-Hypacrosaurus-Lambeosaurus, en partie parce que la crête de Parasaurolophus est dépourvue du fin » coxcomb » osseux qui constitue la partie supérieure de la crête des trois autres lambéosaurins.
Hypothèses rejetées concernant la fonction
Plusieurs des premières suggestions se concentraient sur les adaptations à un mode de vie aquatique, suivant l’hypothèse que les hadrosauridés étaient amphibies, une ligne de pensée commune jusque dans les années 1960. Ainsi, Alfred Sherwood Romer a proposé qu’il servait de tuba, Martin Wilfarth qu’il s’agissait d’un accessoire pour un proboscis mobile utilisé comme tube respiratoire ou pour la collecte de nourriture, Charles M. Sternberg qu’il servait de trappe à air pour empêcher l’eau d’entrer dans les poumons, et Ned Colbert qu’il servait de réservoir d’air pour les séjours prolongés sous l’eau.
D’autres propositions étaient de nature plus physique. Comme mentionné ci-dessus, William Parks a suggéré qu’il était relié aux vertèbres par des ligaments ou des muscles, et qu’il aidait à déplacer et à soutenir la tête. Othenio Abel a proposé qu’elle soit utilisée comme une arme dans les combats entre membres de la même espèce, et Andrew Milner a suggéré qu’elle puisse être utilisée comme un déflecteur de feuillage, comme la crête du casque (appelée « casque ») du casoar. D’autres propositions encore font du logement d’organes spécialisés la fonction principale. Halszka Osmólska a suggéré qu’elle abritait des glandes à sel, et John Ostrom a suggéré qu’elle abritait des zones étendues pour le tissu olfactif et un sens de l’odorat bien amélioré des lambeosaurines, qui n’avaient pas de capacités défensives évidentes. Une suggestion inhabituelle, faite par le créationniste Duane Gish, est que la crête abritait des glandes chimiques qui lui permettaient de lancer des jets de « feu » chimique sur les ennemis, comme le scarabée bombardier des temps modernes.
La plupart de ces hypothèses ont été discréditées ou rejetées. Par exemple, il n’y a pas de trou à l’extrémité de la crête pour une fonction de plongée en apnée. Il n’y a pas de cicatrices musculaires pour un proboscis et il est douteux qu’un animal avec un bec en ait besoin. En tant que sas proposé, il n’aurait pas empêché l’eau d’entrer. Le réservoir d’air proposé aurait été insuffisant pour un animal de la taille de Parasaurolophus. D’autres hadrosauridés avaient de grosses têtes sans avoir besoin de grandes crêtes creuses pour servir de points d’attache à des ligaments de soutien. De plus, aucune des propositions n’explique pourquoi la crête a une telle forme, pourquoi d’autres lambeosaurinés devraient avoir des crêtes d’apparence très différente mais remplissant une fonction similaire, comment des hadrosauridés sans crête ou avec une crête pleine pouvaient se passer de telles capacités, ou pourquoi certains hadrosauridés avaient des crêtes pleines. Ces considérations ont un impact particulier sur les hypothèses basées sur l’augmentation des capacités des systèmes déjà présents chez l’animal, comme les hypothèses de la glande saline et de l’olfaction, et indiquent que ce n’étaient pas des fonctions primaires de la crête. De plus, les travaux sur la cavité nasale des lambeosaurinés montrent que les nerfs olfactifs et les tissus sensoriels correspondants se trouvaient en grande partie à l’extérieur de la portion des voies nasales de la crête, de sorte que l’expansion de la crête avait peu à voir avec l’odorat.
Restauration de vie de Parasaurolophus walkeri avec une hypothétique collerette de peau et des rayures sur sa crête.
Au lieu de cela, les fonctions sociales et physiologiques sont devenues plus soutenues comme fonction(s) de la crête, se concentrant sur l’identification visuelle et auditive et la communication. En tant que grand objet, la crête a une valeur évidente en tant que signal visuel, et distingue cet animal de ses contemporains. La grande taille des orbites des hadrosauridés et la présence d’anneaux sclérotiques dans les yeux impliquent une vision aiguë et des habitudes diurnes, preuve que la vue était importante pour ces animaux. Si, comme cela est couramment illustré, une collerette de peau s’étendait de la crête au cou ou au dos, la présentation visuelle proposée aurait été beaucoup plus voyante. Comme le suggèrent d’autres crânes de lambeosaurins, la crête de Parasaurolophus permettait probablement à la fois l’identification de l’espèce (comme la séparation avec Corythosaurus ou Lambeosaurus) et la détermination entre les mâles et les femelles, sur la base de la forme et de la taille.
Fonction sonore
Modèle informatique des passages nasaux dans la crête de P. tubicen.
Cependant, l’apparence externe de la crête ne correspond pas à l’anatomie interne complexe des passages nasaux, ce qui suggère une autre fonction comptabilisée pour l’utilisation de l’espace interne. Carl Wiman a été le premier à proposer, en 1931, que les passages avaient une fonction de signalisation auditive, comme un cromorne ; Hopson et David B. Weishampel ont repris cette idée dans les années 1970 et 1980. Hopson a découvert qu’il existe des preuves anatomiques que les hadrosauridés avaient une forte capacité auditive. Il existe au moins un exemple, chez le Corythosaurus apparenté, d’un étrier mince (os de l’oreille reptilien) en place, qui, combiné à un grand espace pour le tympan, implique une oreille moyenne sensible. En outre, le lagena de l’hadrosauridé est allongé comme celui d’un crocodilien, ce qui indique que la partie auditive de l’oreille interne était bien développée. Selon Weishampel, P. walkeri était capable de produire des fréquences de 48 à 240 Hz, et P. cyrtocristatus (interprété comme une forme de crête juvénile) de 75 à 375 Hz. En se basant sur la similarité des oreilles internes des hadrosauridés avec celles des crocodiles, il a également proposé que les hadrosauridés adultes soient sensibles aux hautes fréquences, telles que celles que leur progéniture pourrait produire. Selon Weishampel, cela correspond à une communication entre parents et progéniture.
La modélisation informatique d’un spécimen bien conservé de P. tubicen, dont les passages d’air sont plus complexes que ceux de P. walkeri, a permis de reconstituer le son possible que produisait sa crête. Le trajet principal résonne à environ 30 Hz, mais l’anatomie compliquée du sinus provoque des pics et des vallées dans le son.
Fonction de refroidissement
La grande surface et la vascularisation de la crête suggèrent également une fonction thermorégulatrice. P.E. Wheeler a suggéré pour la première fois cette utilisation en 1978 comme un moyen de garder le cerveau au frais. Teresa Maryańska et Osmólska ont également proposé la thermorégulation à peu près au même moment, et Sullivan et Williamson s’y sont intéressés plus avant. La discussion de David Evans en 2006 sur les fonctions de la crête des lambéosaurins était favorable à l’idée, au moins comme facteur initial de l’évolution de l’expansion de la crête.
Dans les médias
Parasaurolophus est un dinosaure très populaire. Il était dans le film Disney’s Dinosaur en tant que membre du troupeau. Il a également fait plusieurs apparitions dans le célèbre documentaire Clash of the Dinosaurs. Il est également apparu dans la populaire émission de télévision Prehistoric Park, où il est devenu la proie des grands carnivores Deinosuchus et Albertosaurus. Il a fait quelques apparitions dans les films Jurassic Park, en tant que membre du troupeau dans le premier film, puis retenu en captivité par des chasseurs dans le deuxième et courant avec le Corythosaurus dans le troisième. et le personnage Dweeb dans « We’re Back ! A Dinosaur’s Story », est lui-même un Parasaurolophus.
Le Parasaurolophus apparaît aussi dans Turok, comme un mangeur de plantes docile qui n’est normalement pas nuisible, mais qui peut être agressif s’il est sévèrement provoqué.
Il y a aussi un zord Parasaurolophus dans Power Rangers : Dino Thunder et Power Rangers : Dino Charge.
Parasaurolophus apparaît brièvement au début du film de Disney Pixar Le Bon Dinosaure.
Voir aussi
Parasaurolophus/Galerie
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, page 229. Publié : 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs : The Encyclopedia, chapitre : Parasaurolophus, pages 678-684. Publié : 1997, McFarland & Co, à Jefferson, Caroline du Nord. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull et Wright, Hadrosaurian Dinosaurs of North America, pages 209-213.
- Brett-Surman, Michael K. et Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks : Dinosaures cératopsiens et ornithopodes, chapitre : Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, pages 135-169. Publié, 2006, Indiana University Press, à Bloomington et Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B. ; et Forster, Catherine A, Weishampel, David B. ; Osmólska, Halszka ; et Dodson, Peter (eds.) The Dinosauria, 2e édition, chapitre : Hadrosauridae, pages 438-463. Publié : 2004, University of California Press, à Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, un nouveau genre et une nouvelle espèce de dinosaure trachodonte à crête, volume 13, pages 1-32. Publié : 1922, Études de l’Université de Toronto, série géologie.
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. et Williamson, Thomas E. Un nouveau crâne de Parasaurolophus (Dinosauria : Hadrosauridae) de la formation Kirtland du Nouveau-Mexique et une révision du genre, de la série New Mexico Museum of Natural History and Science Bulletin, 15, pages 1-52. Publié : 1999, Musée d’histoire naturelle et des sciences du Nouveau-Mexique, à Albuqueque, Nouveau-Mexique.
- 8.0 8.1 8.2 Ostrom, John H. 1961 Une nouvelle espèce de dinosaure hadrosaurien du Crétacé du Nouveau-Mexique, Journal of Paleontology, volume 35, 3e numéro, aux pages 575-577.
- Gilmore, Charles W., Sur le genre Stephanosaurus, avec une description du spécimen type de Lambeosaurus lambei, volume 38, numéro 43, pages 29-48, Parcs. Publié : 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. and Horner, Jack R., Weishampel, David B. ; Osmólska, Halszka ; and Dodson, Peter (eds.) The Dinosauria, 1st edition, Chapter : Hadrosauridae, pages 534-561. Publié : 1990, University of California Press à Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., et Reisz, Robert R. 2007. Anatomie et relations de Lambeosaurus magnicristatus, un dinosaure hadrosauridé à crête (Ornithischia) de la formation Dinosaur Park, Alberta, du Journal of Vertebrate Paleontology, vol. 27 numéro 2, aux pages 373-393.
- Godefroit, Pascal, Shuqin Zan ; et Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., un dinosaure lambéosaurin du Maastrichtien supérieur du nord-est de la Chine, tiré du Compte Rendus de l’Académie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, pages 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Revue des Hadrosauridae (Dinosauria, Ornithischia) du bassin de San Juan, Nouveau-Mexique Lucas, S.G. ; et Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Publié par le New Mexico Museum of Natural History and Science, à Albuqueque, Nouveau-Mexique. Pages 191-213.
- Liddell, Henry George et Robert Scott, 1980. Un lexique grec-anglais (édition abrégée) Publié : Oxford University Press au Royaume-Uni. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. et Evans, David C., Currie, Phillip J., et Koppelhus, Eva (eds.). Dinosaur Provincial Park : A Spectacular Ancient Ecosystem Revealed, Chapitre : Ornithischian Dinosaurs. 2005, publié : Indiana University Press, à Bloomington. Pages 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R. ; et Dupuis, Kevin, 2007. Une boîte crânienne juvénile de Parasaurolophus provenant du parc provincial Dinosaur, Alberta, avec des commentaires sur l’ontogenèse des crêtes dans le genre. Journal of Vertebrate Paleontology, vol. 27, numéro 3 pages 642-650.
- Currie, Phillip J ; et Russell, Dale A. « Distribution géographique et stratigraphique des restes de dinosaures » dans Dinosaur Provincial Park, p. 553.
- Weishampel, David B. (1990). « Distribution des dinosaures », dans The Dinosauria (1ère), p. 63-139.
- 19.0 19.1 19.2 Weishampel, David B. ; Barrett, Paul M. ; Coria, Rodolfo A. ; Le Loeuff, Jean ; Xu Xing ; Zhao Xijin ; Sahni, Ashok ; Gomani, Elizabeth, M.P. ; et Noto, Christopher R. (2004). » Dinosaur Distribution « , dans The Dinosauria (2e), p. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide au Nouveau-Mexique, de la Nova Acta Regia Societas Scientarum Upsaliensis, série 4, vol. 7, numéro 5. (allemand). Pages 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, publié par Cassell Ltd. 1979, édition 5, à Londres. ISBN 0-304-52257-0 Page 883.
- Weishampel, David B. et Jensen, James A. 1979. Parasaurolophus (Reptilia : Hadrosauridae) de l’Utah, du Journal of Paleontology, vol. 53, numéro 6, pages 1422-1427.
- Eberth, David A. 2005. » La géologie « , dans Dinosaur Provincial Park, p. 54-82.
- Braman, Dennis R., et Koppelhus, Eva B. 2005. » Campanian palynomorphs « , dans Dinosaur Provincial Park, p. 101-130.
- Russell, Dale A. An Odyssey in Time : Dinosaures d’Amérique du Nord, 1989. Éditeur : NorthWord Press, à Minocqua, Wisconsin. ISBN 1-55971-038-1 Pages 160-164.
- Bakker, Robert T. 1986. The Dinosaur Heresies : New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, publié par William Morrow, à New York. ISBN 0-8217-2859-8 Page 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs : Un aperçu original et convaincant de la vie dans le royaume des dinosaures, chapitre : Hadrosauridés II. 1985. Publié par Crescent Books, à New York. Pages 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Homologies de la cavité nasale et fonction de la crête crânienne chez les dinosaures lambeosaurins, du Journal of Paleobiology, vol. 32, numéro 1, pages 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. L’évolution des structures d’affichage crânien chez les dinosaures hadrosauriens , du Journal of Paleobiology, volume un, numéro un, pages 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology , de l’Université de Chicago Press, à la page 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, du Journal d’Orion (Munich), vol. 2. pp. 525-532 (langue allemande).
- Sternberg, Charles M. 1935. Hadrosaures à capuchon de la série de Belly River du Crétacé supérieur du Journal du ministère des Mines du Canada Bulletin (Série géologique), volume 77, numéro 52, aux pages 1-37.
- Colbert, Edwin H. The Dinosaur Book : The Ruling Reptiles and their Relatives, publié en 1945 par le Musée américain d’histoire naturelle, Man and Nature Publications, 14, à New York. Page 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas du Journal de Jarbuch Naturwissenschaften, volume 12, numéro 36, aux pages 709-716. (Allemand) 1924.
- Osmólska, Halszka, 1979. Glandes salines nasales chez les dinosaures, du Journal de Acta Palaeontologica Polonica, vol. 24, pages 205-215.
- 36,0 36,1 Ostrom, John H., 1962. Les crêtes crâniennes des dinosaures hadrosauriens, tiré du Journal of Postilla, vol. 62, pages 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, publié par Master Books, à Green Forest. ISBN 0-89051-165-9 Page 82.
- 38,0 38,1 Weishampel, David B., 1981. Analyses acoustiques de la vocalisation potentielle chez les dinosaures lambeosaurins (Reptilia:Ornithischia), du Journal of Paleobiology, vol. 7, numéro 2, p. 252-261.
- Weishampel David B., 1981. La cavité nasale des hadrosauridés lambeosaurins (Reptilia:Ornithischia) : anatomie comparative et homologies, du Journal of Paleontology, vol. 55, numéro 5, pp. 1046-1057.
- Les scientifiques utilisent la paléontologie numérique pour produire la voix du dinosaure Parasaurolophus, par les Sandia National Laboratories (1997-12-05). Récupéré le 20 janvier 2009.
- Diegert, Carl F. et Williamson, Thomas E., 1998. Un modèle acoustique numérique de l’hadrosaure lambeosaurin Parasaurolophus tubicen du Journal of Vertebrate Paleontology, vol. 18, numéro 3, Suppl. p. 38A.
- Sullivan, Robert M. et Williamson, Thomas E., 1996. Un nouveau crâne de Parasaurolophus (forme à longue crête) du Nouveau-Mexique : caractéristiques externes et internes (tomodensitométriques) et leurs implications fonctionnelles, du Journal of Vertebrate Paleontology, vol. 16, numéro 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Structure de refroidissement élaborée du SNC chez les grands dinosaures Journal of Nature, vol. 275, on pp. 441-443.
- Maryańska, Teresa et Osmólska, Halszka, en 1979. Aspects de l’anatomie crânienne des hadrosauriens, du Journal of Lethaia, vol. 12, sur les pp. 265-273.
- Les scientifiques utilisent la paléontologie numérique pour produire la voix du dinosaure Parasaurolophus ; de Sandia National Laboratories.
- Restaurations de P. walkeri et P. cyrtocristatus, à la même échelle, par Scott Hartman ; chez Skeletal Drawing.com.
- Parasaurolophus page sur Enchanted Learning.com
.