














Név: Párducaspelaea.
Fonetikai: Pán-fee-rah lee-oh spe-lay-ah.
Név: Pán-fee-rah lee-oh spe-lay-ah: Georg August Goldfuss – 1810.
Besorolás: Chordata, Mammalia, Carnivora,Felidae, Panthera.
Fajok: P.spelaea.
Táplálkozás:
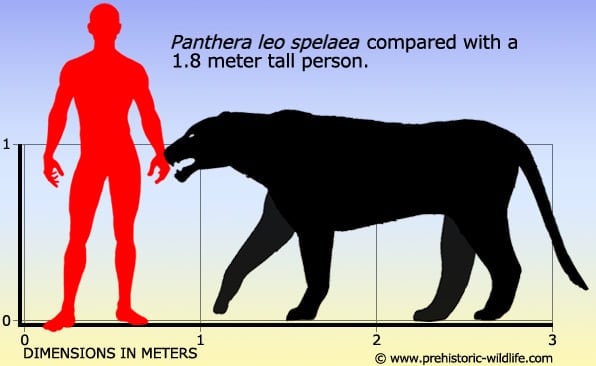
Méret: 1,2 méter magas a vállnál, 2,1 méter hosszú, de egyes maradványok ennél valamivel nagyobb méretre utalnak.
Elismert lelőhelyek: Egész Eurázsiában.
Időszak: Néha megjegyzik, hogy a holocénben is fennmaradt, körülbelül Kr.u. 1-ig.
Fosszilis képviselet:
Közeli rokonához, az amerikai oroszlánhoz (Pantheraatrox) hasonlóan az eurázsiai barlangi oroszlán jelenleg vitatott helyet foglal el a Panthera nemzetség többi tagja között.Az eurázsiai barlangi oroszlánt egykor önálló fajnak tekintették, végül szélesebb körben az afrikai oroszlán (Panthera leo) alfajaként kezelték, azonban egy 2016-os tanulmány (Barnett et al.) és egy 2020-as tanulmány (Stanton et al.) mára újra önálló fajként határozta meg az európai barlangi oroszlánt.
Az eurázsiai barlangi oroszlán vitatott helye ellenére a paleontológusok hajlamosak egyetérteni abban, hogy az idősebb Pantheraleo fossilisből fejlődött ki.Érdekes módon a Panthera leo fossilis valójában nagyobb volt, mint az európai barlangi oroszlán (bár egy kicsivel kisebb, mint az amerikai oroszlán). Általában az állatok az egymást követő generációkkal nagyobbak lesznek, kivéve, ha ökológiai tényezők játszanak közre, mint például a faj számára rendelkezésre álló táplálék vagy zsákmány csökkenése.
Az eurázsiai barlangi oroszlán elterjedtebb nevét a barlangokban talált maradványainak nagy számáról kapta. Az eredmény azonban valójában egy kicsit elnevezése, mivel az eurázsiai barlangi oroszlán más helyekről is ismert, és úgy tűnik, hogy nagyon jól tűrte a hideget, amíg volt elegendő zsákmány a vadászathoz.Ugyanakkor barlangokba is behatoltak, és úgy gondolják, hogy ezt azért tehették, hogy ellopják a barlangi medvebocsokat (Ursusspelaeus), valamint hogy gyenge téli álmot alvó egyedekkel táplálkozzanak.
Az eurázsiai barlangi oroszlán kollagénjének izotópelemzése alátámasztja azt az elképzelést, hogy legalábbis egyes populációk rendszeresen ettek fiatal barlangi medvéket, valamint nagy mennyiségű rénszarvast. Az izotópelemzés azon az elven alapul, hogy a növényevő állatok (a barlangi medvéről úgy gondolják, hogy elsősorban növényevő, esetenként mindenevő) felvesznek bizonyos izotópokat, amelyek attól függően változnak, hogy milyen növényeket esznek, és ezt az izotópot viszont továbbadják az őket fogyasztó ragadozóknak.Bizonyos izotópok nagyobb mennyiségét bizonyos állatokra lehet visszavezetni, ami egy esetleges zsákmányszerzésre utal. Ez a ragadozás lehet az oka annak, hogy olyan sok oroszlánmaradványt találtak barlangokban, mivel elkerülhetetlen, hogy a barlangi medvék egy része visszavágjon, beleértve az álmukból felébresztetteket és a kölykeiket védő anyamedvéket is.
Az eurázsiai barlangi oroszlán nem az egyetlen nagyragadozó volt Eurázsia északi részén, hiszen a Homotherium, a szurikátafogú macska és a barlangi hiéna is aktívan vadászott zsákmányra.Azonban mind a Homotherium, mind a Cavehyena a nyílt síkságok vadászai voltak, és úgy tűnik, hogy előszeretettel vadásztak olyan nagytestű zsákmányokra, mint a gyapjas mamutok és a gyapjas orrszarvúak (például a Coelodonta).Az eurázsiai barlangi oroszlán azonban úgy tűnik, hogy sűrűbben borított területeken vadászott, például erdőkben, amelyeket szarvasok népesítettek be, és amelyek nagyobb mennyiségű fedezékkel rendelkeztek, ami lehetővé tette számukra a lesből támadás taktikáját. Így, bár ezek a ragadozók egy időben voltak aktívak ezeken a kontinenseken, különböző ökoszisztémákban működtek, ami csökkentette az egymás közötti versenyt.
Míg a nagyemlős ragadozók többsége úgy tűnik, hogy a pleisztocén végén a nagy zsákmányállatok hirtelen eltűnésével eltűnt, a barlangi oroszlánról nehezebb bizonyosságot szerezni. Egyes bizonyítékok arra utalnak, hogy Délkelet-Európában kis populációkban még majdnem tízezer évig éltek, bár az ismert maradványok többsége nem nyúlik túl a pleisztocénen.Egy másik érv a késői túlélés mellett az, hogy ha a rénszarvas valóban nagy részét képezte a barlangi oroszlánok étrendjének, akkor a zsákmányforrásuk nem tűnt el.Ezzel szemben azonban más élőlények, például a barlangi medvék csökkenése arra utal, hogy a barlangi oroszlánok csak kisebb populációkban létezhettek.
Az eurázsiai barlangi oroszlánok végleges eltűnésének tényleges időpontjától függetlenül, végső pusztulásuk valószínűleg az új ragadozókkal, különösen a farkasokkal és a korai emberekkel való fokozott verseny miatt következett be.A pleisztocén végével a nyílt síkságok nagy részét erdők váltották fel, amelyekhez a meglévőmegafauna nem alkalmazkodott, ami eltűnésükhöz vezetett.Úgy tűnik, hogy az eurázsiai barlangi oroszlánok otthonosan mozogtak ebben a környezetben, így a változásnak alkalmasnak kellett lennie arra, hogy sikeresebbek legyenek. A farkasok azonban szintén alkalmasak voltak erre az élőhelyre, de korábban a korábbi kiterjedt síkságok miatt korlátozott volt az elterjedésük.Mivel már nem kellett szembenézniük ezekkel az akadályokkal, könnyebben elterjedhettek, és ugyanarra a zsákmányra vadászhattak, amire a barlangi oroszlánok.
Hacsak a préda nem hihetetlenül nagy számban fordul elő, két ragadozó nem élhet együtt ugyanazon zsákmányra vadászva ugyanabban az ökoszisztémában, és végül az egyik feladja. Míg az eurázsiai barlangi oroszlán sokkal nagyobb és erősebb ragadozó volt, a farkasoknak nem kell annyi táplálék a kisebb testükhöz, ráadásul a farkasok teljesen más vadászati taktikát alkalmaznak a táplálékszerzéshez.Az oroszlánok lesből vadásznak, mert nem tudnak lehagyni egy gyors állatot, például egy szarvast egy egyenes versenyben, egyszerűen nem erre lettek tervezve, mivel aránylag rövidebb lábaik jobban gyorsulnak, de a végsebességük a lábuk lépésének korlátai miatt alacsonyabb sebességnél van korlátozva. Így az oroszlánok vadászati viselkedése inkább az energiatakarékosságról szól, míg a farkasok az energiafelhasználásról. Egy farkasfalka szándékosan kényszerít egy szarvascsordát futásra, hogy kiválaszthassák a lassabb és gyengébb egyedeket.Ezután számukat és nagyobb állóképességüket kihasználva addig zaklatják a célpontjukat, amíg az túl gyenge nem lesz a folytatáshoz. Mivel a farkasok ilyen energetikai taktikára számítanak, megengedhetik maguknak, hogy vakmerőek legyenek, és még mindig nagyobb nyereséget érnek el, ha sikerrel járnak, mindkettő olyan előny, amellyel a barlangi oroszlán nem rendelkezett.
A ragadozói egyenlet harmadik eleme az emberi vadászok, akik szintén ugyanazokra az állatokra vadásztak, amelyekre a barlangi oroszlánok és a farkasok egyaránt vadásznak.Az emberi vadászok rendelkeztek a legjobb előnyökkel, amelyek közé tartoztak a testre szabott fegyverek, a használatukhoz szükséges intelligencia, és legfőképpen a különböző problémákhoz való alkalmazkodóképesség. A barlangi oroszlánok szerepeltetése a barlangi művészetben bizonyítja, hogy a korai emberek kapcsolatba kerültek velük, és akárcsak a mai Afrikában, ez a kapcsolat időnként élet-halál harc lehetett az emberek és az oroszlánok között.Fizikai képességeit tekintve egy oroszlán könnyen megölhet egy fegyvertelen embert, de ha több fegyveres és együtt dolgozó ember van jelen, az oroszlánnak nincs esélye.
Az, hogy mennyi konfliktus volt az eurázsiai barlangi oroszlán és a korai ember között, továbbra is ellentmondásos téma, mivel az oroszlánok és az emberek közös fosszíliái többféleképpen is értelmezhetők. Úgy tűnik azonban, hogy a korai emberek nagyra becsülték a barlangi oroszlánokat, mivel szerepelnek a barlangi művészetben, és a barlangi medvéhez hasonlóan a korai életmód részei lehettek.A korai embereknek köszönhető az is, hogy egy kicsit többet tudunk az eurázsiai barlangi oroszlán életkori sajátosságairól, amit lehetetlen lenne csak a csontokból megállapítani. Ezek közé a jellemzők közé tartozik a sörény jelenléte a feltehetően hímeknél, a farkak végén lévő tincsek és a kerek, bolyhos fülek, ez utóbbi valószínűleg alkalmazkodás volt, hogy megvédje ezeket a végtagokat a hideg körülményektől.A műalkotást úgy is értelmezték, mintha halvány csíkok futnának végig a testen, ami valójában jó alkalmazkodás lett volna, mivel a barlangi oroszlánok valószínűleg inkább a fákat és bokrokat használták fedezékként, minthogy afrikai rokonaikhoz hasonlóan a hosszú fűben bujkáljanak.Emellett a barlangrajzokon több oroszlán együtt vadászik, ami arra utal, hogy a műalkotásokat készítő emberek a barlangi oroszlánokat falkában figyelték meg.

Pantheraatrox, Pantheraleo fossilis, Pantheraspelaea.
További olvasmányok
– The Pleistocene cave lion, Panthera spelaea(Carnivora, Felidae) fromYakutia, Russia. – Cranium 18, 7-24. – G. F. Baryshnikov & G.Boeskorov – 2001.
– Molecular phylogeny of the extinct cave lion Panthera leospelaea -Molecular Phylogenetics and EvolutionVolume 30, Issue 3 – JoachimBurgera, Wilfried Rosendahl, Odile Loreillea, Helmut Hemmer, TorstenEriksson, Anders Götherström, Jennifer Hiller, Matthew J. Collins,Timothy Wessg & Kurt W. Alt – 2004.
– Evolution of the mane and group-living in the lion (Pantheraleo): areview – Journal of Zoology Volume 263, Issue 4, pages 329-342. -Nobuyuki Yamaguchi, Alan Cooper, Lars Werdelin & David W.Macdonald- 2004.
– Upper Pleistocene Panthera leo spelaea (Goldfuss,1810) skeletonremains from Praha-Podbaba and other lion finds from loess and riverterrace sites in Central Bohemia (Czech Republic). – Bulletin ofGeosciences 82 (2) – Cajus G. Diedrich – 2007.
– Phylogeny of the great cats (Felidae: Pantherinae), and the influenceofossil taxa and missing characters. – Cladistics Vol.24, No.6,pp.977-992. – Per Christiansen – 2008.
– Felső pleisztocén Panthera leo spelaea (Goldfuss,1810) maradványok a Bilstein-barlangokból (Sauerlandi karszt) és hozzájárulás a steppelion taphonomiához, paleobiológiához és szexuális dimorfizmushoz – Annales dePaléontologie 95. kötet, 3. szám – Cajus G. Diedrich – 2009.
– Az oroszlánok (Panthera leo ssp.) filogeográfiája három különböző rendszertant és a genetikai diverzitás késő pleisztocén kori csökkenését mutatja. – MolecularEcology 18 (8): 1668-1677 – Ross Barnett, Beth Shapiro, Ian Barnes,Simony W. Ho, Joachim Burger, Nobuyuki Yamaguchi, Thomas F. G. Highham,H. Todd Wheeler, Wilfred Rosendahl, Andrei V. Sher, Marina Sotnikova,tatiana Kuznetsova, Gennady F. Baryshnikov, Larry D. Martin, C. RichardBurns & Alan Cooper – 2009.
– Isotopic evidence for dietary ecology of cave lion (Pantheraspelaea)in North-Western Europe: Prey choice, competition and implications forextinction. – Quaternary International 245 (2): 249-261. – HervéBocherens, Dorothée G. Drucker, Dominique Bonjean, Anne Bridault,Nicholas J. Conard, Christophe Cupillard, Mietje Germonpré, MarkusHöneisen, Susanne C. Münzel, Hannes Napierala, Marylène Patou-Mathis,Elisabeth Stephan, Hans-Peter Uerpmann, Reinhard Ziegler – 2011.
– Late Pleistocene steppe lion Panthera leo spelaea(Goldfuss, 1810)footprints and bone records from open air sites in northern Germany -Evidence of hyena-lion antagonism and scavenging in Europe – QuaternaryScience ReviewsVolume 30, Issues 15-16 – Cajus G. Diedrich – 2011.
– A legnagyobb európai oroszlán Panthera leo spelaea(Goldfuss 1810)populáció a németországi Zoolithen-barlangból: Európa specializált barlangi medvepredátorai – Történeti biológia: An International Journal ofPaleobiology Volume 23, Issue 2-3 Cajus G. Diedrich – 2011.
– Late Pleistocene steppe lion Panthera leo spelaea(Goldfuss 1810)skeleton remains of the Upper Rhine Valley (SW Germany) andcontributions to their sexual dimorphism, taphonomy and habitus -Historical Biology: An International Journal of Paleobiology vol 24,issue 1. – Cajus G. Diedrich & Thomas Rathgeber – 2011.
– Palaeopulations of Late Pleistocene Top Predators in Europe: IceAge Spotted Hyenas and Steppe Lions in Battle and Competition aboutPrey. – Paleontology Journal. 2014: 1-34. – C. G. Diedrich – 2014.
– Population Demography and Genetic Diversity in the Pleistocene CaveLion. – Open Quaternary. 1 (1): Art. 4. – E. Ersmark, L. Orlando, E.Sandoval-Castellanos, I. Barnes, R. Barnett, A. Stuart, A. Lister& L. Dalén – 2015.
– On the discovery of a barlangi oroszlán a Malyi Anyui folyóból (Chukotka,Oroszország). – Quaternary Science Reviews. 117: 135-151. – I. Kirillova, A.V. Tiunov, V. A. Levchenko, O. F. Chernova, V. G. Yudin, F. Bertuch& F. K. Shidlovskiy – 2015.
– Mitogenomics of the Extinct Cave Lion, Panthera spelaea(Goldfuss,1810), Resolve its Position within the Panthera Cats. – OpenQuaternary. 2: 4. – Ross Barnett, Marie Lisandra Zepeda, Mendoza, AndréElias Rodrigues Soares, Simon Y W Ho, Grant Zazula, Nobuyuki Yamaguchi,Beth Shapiro, Irina V Kirillova, Greger Larson, M Thomas & PGilbert – 2016.
– Egy barlangi oroszlán (Panthera spelaea Goldfuss,1810) szőrének morfológiai és genetikai azonosítása és izotópos vizsgálata a MalyiAnyui folyóból (Csukotka, Oroszország). – Quaternary Science Reviews. 142:61-73. – O. F. Chernova, I. V. Kirillova, B. Shapiro, F. K.Shidlovskiy, A. E. R. Soares, V. A. Levchenko & F. Bertuch -2016.
– Under the Skin of a Lion: Unique Evidence of Upper PaleolithicExploitation and Use of Cave Lion (Panthera spelaea)from the LowerGallery of La Garma (Spain). – PLOS ONE. 11 (10): e0163591. – M. Cueto,E. Camarós, P. Castaños, R. Ontañón & P. Arias – 2017.
– Early Pleistocene origin and extensive intra-species diversity of theextinct cave lion. – Scientific Reports. 10: 12621. – David W. G.Stanton, Federica Alberti, Valery Plotnikov, Semyon Androsov, SemyonGrigoriev, Sergey Fedorov, Pavel Kosintsev, Doris Nagel, SergeyVartanyan, Ian Barnes, Ross Barnett, Erik Ersmark, Doris Döppes, MietjeGermonpré, Michael Hofreiter, Wilfried Rosendahl, Pontus Skoglund& Love Dalén – 2020.
—————————————————————————-
Véletlenszerű kedvencek
 |
 |
 |
 |
Adatvédelem& Sütikre vonatkozó irányelvek
.