

Parasaurolophus (pronuncia /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, comunemente anche /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; che significa “lucertola crestata vicina” in riferimento a Saurolophus) è un genere di dinosauro ornitopode del tardo Cretaceo di quello che oggi è il Nord America, circa 76-73 milioni di anni fa. Era un erbivoro che camminava sia come bipede che come quadrupede. Sono riconosciute tre specie: P. walkeri (la specie tipo), P. tubicen, e il P. cyrtocristatus a cresta corta. Resti sono noti da Alberta, Canada, e New Mexico e Utah, USA. Fu descritto per la prima volta nel 1922 da William Parks da un cranio e uno scheletro parziale in Alberta.
Parasaurolophus è un adrosauride, parte di una diversa famiglia di dinosauri del Cretaceo noti per la loro gamma di bizzarri ornamenti della testa. Questo genere è noto per la sua grande ed elaborata cresta cranica, che al suo massimo forma un lungo tubo curvo che sporge verso l’alto e indietro dal cranio. Charonosaurus dalla Cina, che potrebbe essere stato il suo parente più prossimo, aveva un cranio simile e potenzialmente una cresta simile. La cresta è stata molto discussa dagli scienziati; il consenso è che le funzioni principali includevano il riconoscimento visivo sia della specie che del sesso, la risonanza acustica e la termoregolazione. È uno degli ornitorinchi più rari, essendo conosciuto solo da una manciata di buoni esemplari.
Parasaurolophus è tra i dinosauri più ben riconosciuti, ed è frequentemente ritratto in spettacoli televisivi, film, libri, merchandising e altri media legati ai dinosauri. Alice Perry
Descrizione

Le dimensioni del Parasaurolophus walkeri rispetto ad un uomo.
Come la maggior parte dei dinosauri, lo scheletro del Parasaurolophus è incompletamente conosciuto. La lunghezza dell’esemplare tipo di P. walkeri è stimata in 9,5 metri (31 piedi). Il suo cranio è lungo circa 1,6 metri (5,2 piedi), compresa la cresta, mentre il cranio tipo di P. tubicen è lungo oltre 2,0 metri (6,6 piedi), indicando un animale più grande. Il suo peso è stimato in 2,5 tonnellate (2,7 tonnellate). Il suo singolo arto anteriore conosciuto era relativamente corto per un adrosauro, con una scapola corta ma larga. L’osso della coscia misura 103 centimetri di lunghezza in P. walkeri ed è robusto per la sua lunghezza se confrontato con altri adrosauridi. Anche la parte superiore del braccio e le ossa pelviche erano pesantemente costruite.
Come altri adrosauridi, era in grado di camminare sia su due gambe che su quattro. Probabilmente preferiva procacciarsi il cibo su quattro zampe, ma correva su due. Le spine neurali delle vertebre erano alte, come era comune nei lambeosaurini; più alte sopra le anche, aumentavano l’altezza della schiena. Per P. walkeri sono note le impronte della pelle, che mostrano squame uniformi simili a tubercoli, ma nessuna struttura più grande.
Parasaurolophus walkeri con dettaglio delle squame.
La caratteristica più evidente era la cresta cranica, che sporgeva dalla parte posteriore della testa ed era formata dalla premaxilla e dalle ossa nasali. L’esemplare tipo di P. walkeri ha una tacca nelle spine neurali vicino a dove la cresta avrebbe colpito la schiena, ma questa potrebbe essere una patologia peculiare di questo individuo. William Parks, che diede il nome al genere, ipotizzò che un legamento corresse dalla cresta alla tacca per sostenere la testa. Anche se l’idea sembra improbabile, Parasaurolophus è talvolta restaurato con un lembo di pelle dalla cresta al collo.
La cresta era cava, con tubi distinti che portavano da ogni narice alla fine della cresta prima di invertire la direzione e dirigersi di nuovo giù per la cresta e nel cranio. I tubi erano più semplici in P. walkeri, e più complessi in P. tubicen, dove alcuni tubi erano ciechi e altri si incontravano e si separavano. Mentre P. walkeri e P. tubicen avevano lunghe creste con solo una leggera curvatura, P. cyrtocristatus aveva una cresta corta con un profilo più circolare.
Classificazione
Come implica il suo nome, Parasaurolophus fu inizialmente pensato per essere strettamente legato a Saurolophus a causa della sua cresta superficialmente simile. Tuttavia, è stato presto rivalutato come un membro della sottofamiglia lambeosaurina degli adrosauridi-Saurolophus è un adrosaurino. Di solito viene interpretato come un ramo separato dei lambeosaurini, distinto dai Corythosaurus, Hypacrosaurus e Lambeosaurus con la cresta a caschetto. Il suo parente più prossimo conosciuto sembra essere Charonosaurus, un lambeosaurino con un cranio simile (ma senza cresta completa ancora) dalla regione dell’Amur della Cina nord-orientale, e i due possono formare un clade Parasaurolophini. P. cyrtocristatus, con la sua cresta corta e più rotonda, potrebbe essere la più basale delle tre specie conosciute di Parasaurolophus, o potrebbe rappresentare esemplari subadulti o femminili di P. tubicen.
Storia
Ricerca e denominazione
Cast dell’esemplare tipo incompleto di Parasaurolophus walkeri a Warszawa.
Significato “lucertola crestata vicina”, il nome Parasaurolophus deriva dal greco para/παρα “accanto” o “vicino”, saurus/σαυρος “lucertola” e lophos/λοφος “cresta”. Si basa su ROM 768, un cranio e uno scheletro parziale mancante della maggior parte della coda e delle zampe posteriori sotto le ginocchia, che fu trovato da un gruppo di ricercatori dell’Università di Toronto nel 1920 vicino a Sand Creek lungo il fiume Red Deer in Alberta, Canada. Queste rocce sono ora conosciute come la Formazione di Dinosaur Park del Cretaceo superiore di età Campaniana. William Parks chiamò l’esemplare P. walkeri in onore di Sir Byron Edmund Walker, presidente del consiglio di amministrazione del Royal Ontario Museum. I resti di Parasaurolophus sono rari in Alberta, con solo un altro cranio parziale dalla (probabilmente) Formazione Dinosaur Park, e tre esemplari Dinosaur Park privi di cranio, forse appartenenti al genere. In alcune liste faunistiche, c’è una menzione di possibile materiale di P. walkeri nella Formazione Hell Creek del Montana, un’unità rocciosa di età tardo-maastrichtiana. Questa presenza non è stata notata da Sullivan e Williamson nella loro revisione del 1999 del genere, e non è stata ulteriormente elaborata altrove.
Nel 1921, Charles H. Sternberg recuperò un cranio parziale (PMU.R1250) da quella che ora è conosciuta come la formazione Kirtland leggermente più giovane nella contea di San Juan, New Mexico. Questo esemplare fu inviato a Uppsala, Svezia, dove Carl Wiman lo descrisse come una seconda specie, P. tubicen, nel 1931. L’epiteto specifico deriva dal latino tǔbǐcěn “trombettista”. Un secondo cranio quasi completo di P. tubicen (NMMNH P-25100) è stato trovato nel New Mexico nel 1995. Usando la tomografia computerizzata di questo cranio, Robert Sullivan e Thomas Williamson hanno dato al genere un trattamento monografico nel 1999, coprendo aspetti della sua anatomia e tassonomia, e le funzioni della sua cresta. Williamson ha poi pubblicato una revisione indipendente dei resti, in disaccordo con le conclusioni tassonomiche.
John Ostrom ha descritto un altro buon esemplare (FMNH P27393) dal Nuovo Messico come P. cyrtocristatus nel 1961. Include un cranio parziale con una cresta corta e arrotondata, e gran parte dello scheletro postcraniale tranne i piedi, il collo e parti della coda. Il suo nome specifico deriva dal latino curtus “accorciato” e cristatus “crestato”. L’esemplare è stato trovato nella parte superiore della Formazione Fruitland o, più probabilmente, alla base della sovrastante Formazione Kirtland. La gamma di questa specie è stata ampliata nel 1979, quando David B. Weishampel e James A. Jensen hanno descritto un cranio parziale con una cresta simile (BYU 2467) dalla Formazione Kaiparowits di età Campaniana della Contea di Garfield, Utah. Da allora, un altro cranio è stato trovato nello Utah con la morfologia della cresta corta/rotonda di P. cyrtocristatus.
Specie
Calco del cranio di Parasaurolophus walkeri, Natural History Museum, Londra.
Calco del cranio di Parasaurolophus cyrtocristatus, Field Museum of Natural History.
La specie tipo P. walkeri, dall’Alberta, è conosciuta da un solo esemplare. Si differenzia da P. tubicen per avere tubi più semplici nella sua cresta, e da P. cyrtocristatus per avere una cresta lunga e non arrotondata e un braccio superiore più lungo dell’avambraccio.
P. tubicen, dal Nuovo Messico, è conosciuto dai resti di almeno tre individui. È la specie più grande, con passaggi d’aria più complessi nella cresta rispetto a P. walkeri, e una cresta più lunga e dritta di P. cyrtocristatus.
P. cyrtocristatus, dal Nuovo Messico e dallo Utah, è conosciuto da tre possibili esemplari. È la specie più piccola, con una breve cresta arrotondata. Le sue piccole dimensioni e la forma della cresta hanno portato diversi scienziati a suggerire che si tratti di giovani o femmine di P. tubicen, che proviene all’incirca dallo stesso periodo e dalla stessa formazione nel Nuovo Messico. Come notato da Thomas Williamson, il materiale tipo di P. cyrtocristatus è circa il 72% della dimensione di P. tubicen, vicino alla dimensione alla quale altri lambeosaurini sono interpretati per iniziare a mostrare il dimorfismo sessuale definitivo nelle loro creste (~70% della dimensione adulta). Questa posizione è stata respinta in recenti revisioni dei lambeosaurini.
Paleobiologia
Paleoecologia
Gorgosaurus (a sinistra) attacca un Parasaurolophus cyrtocristatus.
Parasaurolophus walkeri, dalla Formazione del Dinosaur Park, era un membro di una fauna diversificata e ben documentata di animali preistorici, inclusi dinosauri ben noti come il Centrosaurus cornuto, Styracosaurus, e Chasmosaurus; gli ornitorinchi Prosaurolophus, Gryposaurus, Corythosaurus, e Lambeosaurus; il tirannosauro Gorgosaurus; e i corazzati Edmontonia ed Euoplocephalus. Era un raro costituente di questa fauna. La Formazione del Parco dei Dinosauri è interpretata come un ambiente a basso rilievo di fiumi e pianure alluvionali che è diventato più paludoso e influenzato da condizioni marine nel corso del tempo, quando la Western Interior Seaway ha trasgredito verso ovest. Il clima era più caldo dell’attuale Alberta, senza gelo, ma con stagioni più umide e più secche. Le conifere erano apparentemente le piante dominanti, con un sottobosco di felci, felci arboree e angiosperme.
Le specie del Nuovo Messico condividevano il loro ambiente con i grandi sauropodi Alamosaurus, Kritosaurus, Pentaceratops cornuto, Nodocephalosaurus corazzato, Saurornitholestes, e tirannosauridi attualmente senza nome. La Formazione Kirtland è interpretata come pianure alluvionali fluviali apparse dopo una ritirata del mare interno occidentale. Le conifere erano le piante dominanti, e i dinosauri cornuti chasmosaurini erano apparentemente più comuni degli adrosauridi.
Alimentazione
Come adrosauride, Parasaurolophus era un grande erbivoro bipede/quadrupede, mangiava le piante con un cranio sofisticato che permetteva un movimento di macinazione analogo alla masticazione. I suoi denti venivano continuamente sostituiti e confezionati in batterie dentarie che contenevano centinaia di denti, di cui solo una manciata relativa era in uso in qualsiasi momento. Usava il suo becco per raccogliere materiale vegetale, che era tenuto nelle mascelle da un organo simile a una guancia. L’alimentazione sarebbe stata da terra fino a circa 4 metri di altezza. Come notato da Bob Bakker, i lambeosaurini hanno becchi più stretti degli adrosaurini, il che implica che Parasaurolophus e i suoi parenti potevano nutrirsi più selettivamente delle loro controparti dal becco largo e senza cresta.
Cresta cranica
Molte ipotesi sono state avanzate su quali funzioni svolgesse la cresta cranica di Parasaurolophus, ma molte sono state screditate. Ora si ritiene che possa aver avuto diverse funzioni: visualizzazione per identificare specie e sesso, amplificazione del suono per la comunicazione e termoregolazione. Non è chiaro quale fosse più significativa in quali momenti dell’evoluzione della cresta e dei suoi passaggi nasali interni.
Differenze tra specie e stadi di crescita
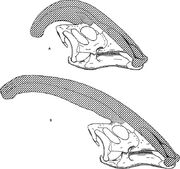
Diagramma che confronta le creste nariali di Parasaurolophus cyrtocristatus (a) e Parasaurolophus walkeri (b).
Come per altri lambeosaurini, si ritiene che la cresta cranica di Parasaurolophus cambiasse con l’età e fosse una caratteristica sessualmente dimorfica negli adulti. James Hopson, uno dei primi ricercatori a descrivere le creste lambeosaurine in termini di tali distinzioni, suggerì che P. cyrtocristatus, con la sua piccola cresta, fosse la forma femminile di P. tubicen. Thomas Williamson suggerì che fosse la forma giovanile. Nessuna delle due ipotesi fu ampiamente accettata. Poiché si conoscono solo sei crani buoni e una scatola cranica giovanile, ulteriore materiale aiuterà a chiarire queste potenziali relazioni. Williamson ha notato che in ogni caso, il Parasaurolophus giovanile probabilmente aveva piccole creste arrotondate come P. cyrtocristatus, che probabilmente crescevano più velocemente quando gli individui si avvicinavano alla maturità. Il recente studio di un caso giovanile precedentemente assegnato a Lambeosaurus, e ora assegnato a Parasaurolophus, fornisce la prova che una piccola cresta tubolare era presente nei giovani. Questo esemplare conserva una piccola svasatura verso l’alto delle ossa frontali che era simile ma più piccola di quella che si vede negli esemplari adulti; negli adulti, le frontali formavano una piattaforma che sosteneva la base della cresta. Questo esemplare indica anche che la crescita della cresta in Parasaurolophus e il profilo facciale degli individui giovani differiva dal modello Corythosaurus-Hypacrosaurus-Lambeosaurus, in parte perché la cresta di Parasaurolophus manca del sottile “coxcomb” osseo che costituisce la parte superiore della cresta degli altri tre lambeosaurini.
Ipotesi rifiutate sulla funzione
Molti dei primi suggerimenti si concentravano sugli adattamenti per uno stile di vita acquatico, seguendo l’ipotesi che gli adrosauridi fossero anfibi, una linea di pensiero comune fino agli anni ’60. Così, Alfred Sherwood Romer propose che servisse come boccaglio, Martin Wilfarth che fosse un attacco per una proboscide mobile usata come tubo per respirare o per raccogliere il cibo, Charles M. Sternberg che servisse come trappola per tenere l’acqua fuori dai polmoni, e Ned Colbert che servisse come riserva d’aria per prolungati soggiorni sott’acqua.
Altre proposte erano di natura più fisica. Come menzionato sopra, William Parks suggerì che fosse unito alle vertebre con legamenti o muscoli, e che aiutasse a muovere e sostenere la testa. Othenio Abel propose che fosse usato come arma in combattimento tra membri della stessa specie, e Andrew Milner suggerì che potesse essere usato come deflettore di fogliame, come la cresta dell’elmo (chiamata ‘casco’) del casuario. Altre proposte facevano dell’alloggiamento di organi specializzati la funzione principale. Halszka Osmólska ha suggerito che ospitasse ghiandole saline, e John Ostrom ha suggerito che ospitasse aree espanse per il tessuto olfattivo e l’olfatto molto migliorato dei lambeosaurini, che non avevano evidenti capacità difensive. Un suggerimento insolito, fatto dal creazionista Duane Gish, è che la cresta ospitasse ghiandole chimiche che gli permettevano di lanciare getti di “fuoco” chimico contro i nemici, simile al moderno scarabeo bombardiere.
La maggior parte di queste ipotesi sono state screditate o respinte. Per esempio, non c’è nessun foro alla fine della cresta per una funzione di snorkeling. Non ci sono cicatrici muscolari per una proboscide ed è dubbio che un animale con un becco ne avrebbe bisogno. Come camera di compensazione proposta, non avrebbe tenuto fuori l’acqua. Il serbatoio d’aria proposto sarebbe stato insufficiente per un animale delle dimensioni di Parasaurolophus. Altri adrosauridi avevano grandi teste senza bisogno di grandi creste cave per servire come punti di attacco per i legamenti di sostegno. Inoltre, nessuna delle proposte spiega perché la cresta ha una tale forma, perché altri lambeosaurini dovrebbero avere creste che sembrano molto diverse ma svolgono una funzione simile, come gli adrosauridi senza cresta o con cresta solida se la cavavano senza tali capacità, o perché alcuni adrosauridi avevano creste solide. Queste considerazioni hanno un impatto particolare sulle ipotesi basate sull’aumento delle capacità di sistemi già presenti nell’animale, come le ipotesi della ghiandola del sale e dell’olfatto, e indicano che queste non erano funzioni primarie della cresta. Inoltre, il lavoro sulla cavità nasale dei lambeosaurini mostra che i nervi olfattivi e il corrispondente tessuto sensoriale erano in gran parte al di fuori della porzione delle fosse nasali nella cresta, quindi l’espansione della cresta aveva poco a che fare con l’olfatto.
Ripristino della vita di Parasaurolophus walkeri con un ipotetico orlo di pelle e strisce sulla cresta.
Invece, le funzioni sociali e fisiologiche sono diventate più sostenute come funzioni della cresta, concentrandosi sull’identificazione e comunicazione visiva e uditiva. Come oggetto di grandi dimensioni, la cresta ha un chiaro valore come segnale visivo, e distingue questo animale dai suoi contemporanei. Le grandi dimensioni delle orbite degli adrosauridi e la presenza di anelli sclerotici negli occhi implicano una visione acuta e abitudini diurne, prova che la vista era importante per questi animali. Se, come è comunemente illustrato, un fronzolo di pelle si estendeva dalla cresta al collo o alla schiena, l’esposizione visiva proposta sarebbe stata molto più vistosa. Come è suggerito da altri crani di lambeosaurini, la cresta di Parasaurolophus permetteva probabilmente sia l’identificazione della specie (come la separazione da Corythosaurus o Lambeosaurus) che la determinazione tra maschi e femmine, sulla base della forma e delle dimensioni.
Funzione sonora
Modello al computer dei passaggi nasali nella cresta di P. tubicen.
Tuttavia, l’aspetto esterno della cresta non corrisponde alla complessa anatomia interna dei passaggi nasali, il che suggerisce un’altra funzione per l’utilizzo dello spazio interno. Carl Wiman fu il primo a proporre, nel 1931, che i passaggi servissero una funzione di segnalazione uditiva, come un crumhorn; Hopson e David B. Weishampel rivisitarono questa idea negli anni ’70 e ’80. Hopson ha scoperto che ci sono prove anatomiche che gli adrosauridi avevano un forte udito. C’è almeno un esempio, nel relativo Corythosaurus, di una sottile staffa (osso dell’orecchio del rettile) al suo posto, che combinato con un grande spazio per un timpano implica un orecchio medio sensibile. Inoltre, la lagena adrosauride è allungata come quella di un coccodrillo, indicando che la porzione uditiva dell’orecchio interno era ben sviluppata. Weishampel ha suggerito che P. walkeri era in grado di produrre frequenze da 48 a 240 Hz, e P. cyrtocristatus (interpretato come una forma giovanile di cresta) da 75 a 375 Hz. Basandosi sulla somiglianza delle orecchie interne degli adrosauridi con quelle dei coccodrilli, ha anche proposto che gli adrosauridi adulti fossero sensibili alle alte frequenze, come quelle che la loro prole potrebbe produrre. Secondo Weishampel, questo è coerente con la comunicazione tra genitori e prole.
La modellazione al computer di un esemplare ben conservato di P. tubicen, con passaggi d’aria più complessi di quelli di P. walkeri, ha permesso la ricostruzione del possibile suono prodotto dalla sua cresta. Il percorso principale risuona a circa 30 Hz, ma la complicata anatomia del seno provoca picchi e valli nel suono.
Funzione di raffreddamento
La grande superficie e la vascolarizzazione della cresta suggeriscono anche una funzione termoregolatrice. P.E. Wheeler ha suggerito per la prima volta questo uso nel 1978 come un modo per mantenere il cervello fresco. Teresa Maryańska e Osmólska hanno anche proposto la termoregolazione più o meno nello stesso periodo, e Sullivan e Williamson si sono interessati ulteriormente. La discussione di David Evans del 2006 sulle funzioni della cresta lambeosaurina era favorevole all’idea, almeno come fattore iniziale per l’evoluzione dell’espansione della cresta.
Nei media
Parasaurolophus è un dinosauro molto popolare. Era nel film Disney’s Dinosaur come membro del branco. Fece anche diverse apparizioni nel famoso documentario Clash of the Dinosaurs. Apparve anche nel popolare show televisivo Prehistoric Park, dove divenne la preda dei grandi carnivori Deinosuchus e Albertosaurus. Fece qualche apparizione nei film di Jurassic Park, come membro del branco nel primo film, per poi essere tenuto prigioniero dai cacciatori nel secondo e correre insieme a Corythosaurus nel terzo. e il personaggio Dweeb in “We’re Back! A Dinosaur’s Story”, è lui stesso un Parasaurolophus.
Il Parasaurolophus appare anche in Turok, come un docile mangiatore di piante che normalmente non è dannoso, ma può essere aggressivo se fortemente provocato.
C’è anche uno zord Parasaurolophus in Power Rangers: Dino Thunder e Power Rangers: Dino Charge.
Parasaurolophus appare brevemente all’inizio del film Disney Pixar The Good Dinosaur.
See Also
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, page 229. Pubblicato: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Chapter: Parasaurolophus, pagine 678-684. Pubblicato: 1997, McFarland & Co, a Jefferson, North Carolina. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull e Wright, Hadrosaurian Dinosaurs of North America, pp. 209-213.
- Brett-Surman, Michael K. e Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, capitolo: Anatomia appendicolare negli adrosauridi nordamericani del Campaniano e del Maastrichtiano, pagine 135-169. Pubblicato, 2006, Indiana University Press, a Bloomington e Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; e Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; e Dodson, Peter (eds.) The Dinosauria, 2nd edition, chapter: Hadrosauridae, pagine 438-463. Pubblicato: 2004, University of California Press, a Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, volume 13, pagine 1-32. Pubblicato: 1922, Università di Toronto Studi, Serie Geologia.
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. e Williamson, Thomas E. Un nuovo cranio di Parasaurolophus (Dinosauria: Hadrosauridae) dalla Formazione Kirtland del Nuovo Messico e una revisione del genere, dalla serie New Mexico Museum of Natural History and Science Bulletin, 15, pagine 1-52. Pubblicato: 1999, New Mexico Museum of Natural History and Science, ad Albuqueque, New Mexico.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, Journal of Paleontology, Volume 35, 3rd issue, on pages 575-577.
- Gilmore, Charles W., Sul genere Stephanosaurus, con una descrizione dell’esemplare tipo di Lambeosaurus lambei, volume 38, numero 43, pagine 29-48, Parks. Pubblicato: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. e Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; e Dodson, Peter (eds.) The Dinosauria, 1st edition, Chapter: Hadrosauridae, pagine 534-561. Pubblicato: 1990, University of California Press a Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., and Reisz, Robert R. 2007. Anatomia e relazioni di Lambeosaurus magnicristatus, un dinosauro adrosauride crestato (Ornithischia) dalla Formazione di Dinosaur Park, Alberta, dal Journal of Vertebrate Paleontology, vol. 27 numero 2, alle pagine 373-393.
- Godefroit, Pascal, Shuqin Zan; e Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., un dinosauro lambeosaurino dal tardo Maastrichtiano della Cina nord-orientale, dal Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, pagine 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Published by New Mexico Museum of Natural History and Science, in Albuqueque, New Mexico. Pagine 191-213.
- Liddell, Henry George e Robert Scott, 1980. A Greek-English Lexicon (Abridged Edition) Pubblicato: Oxford University Press nel Regno Unito. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. e Evans, David C., Currie, Phillip J., e Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, Chapter: Dinosauri ornitischi. 2005, Pubblicato: Indiana University Press, a Bloomington. Pagine 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; e Dupuis, Kevin, 2007. Un caso giovanile di Parasaurolophus dal Dinosaur Provincial Park, Alberta, con commenti sull’ontogenesi della cresta nel genere. Journal of Vertebrate Paleontology, vol. 27, numero 3 pagine 642-650.
- Currie, Phillip J; e Russell, Dale A. “Distribuzione geografica e stratigrafica dei resti di dinosauro” in Dinosaur Provincial Park, p. 553.
- Weishampel, David B. (1990). “Distribuzione dei dinosauri”, in The Dinosauria (1st), pp. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). “Dinosaur Distribution”, in The Dinosauria (2nd), pp. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, dalla Nova Acta Regia Societas Scientarum Upsaliensis, serie 4, vol. 7, numero 5. (Tedesco). Pagine 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, pubblicato da Cassell Ltd. 1979, edizione 5, a Londra. ISBN 0-304-52257-0 Pagina 883.
- Weishampel, David B. e Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) dallo Utah, dal Journal of Paleontology, vol. 53, numero 6, pagine 1422-1427.
- Eberth, David A. 2005. “La geologia”, in Dinosaur Provincial Park, pp. 54-82.
- Braman, Dennis R., e Koppelhus, Eva B. 2005. “Campanian palynomorphs”, in Dinosaur Provincial Park, pp. 101-130.
- Russell, Dale A. An Odyssey in Time: Dinosaurs of North America, 1989. Editore: NorthWord Press, a Minocqua, Wisconsin. ISBN 1-55971-038-1 Pagine 160-164.
- Bakker, Robert T. 1986. Le eresie dei dinosauri: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, pubblicato da William Morrow, a New York. ISBN 0-8217-2859-8 Pagina 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom, capitolo: Hadrosaurids II. 1985. Pubblicato da Crescent Books, a New York. Pagine 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Omologie della cavità nasale e funzione della cresta cranica nei dinosauri lambeosaurini, dal Journal of Paleobiology, vol. 32, issue 1, Pages 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. L’evoluzione delle strutture di visualizzazione del cranio nei dinosauri adrosauriani, dal Journal of Paleobiology, volume uno, numero uno, pagine 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology, dalla University of Chicago Press, a pagina 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, dal Journal of Orion (Monaco), vol. 2. pp. 525-532 (lingua tedesca).
- Sternberg, Charles M. 1935. Adrosauri incappucciati della Belly River Series del Cretaceo superiore dal Journal of the Canada Department of Mines Bulletin (Geological Series), volume 77, issue 52, alle pagine 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, pubblicato nel 1945 dall’American Museum of Natural History, Man and Nature Publications, 14, a New York. Pagina 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas dal Journal of Jarbuch Naturwissenschaften, volume 12, numero 36, alle pagine 709-716. (Tedesco) 1924.
- Osmólska, Halszka, 1979. Ghiandole saline nasali nei dinosauri, dal Journal of Acta Palaeontologica Polonica, vol. 24, pagine 205-215.
- 36.0 36.1 Ostrom, John H., 1962. Le creste craniche dei dinosauri adrosauriani, dal Journal of Postilla, vol. 62, pagine 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, pubblicato da Master Books, in Green Forest. ISBN 0-89051-165-9 Pagina 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), dal Journal of Paleobiology, vol. 7, issue 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, da Journal of Paleontology, vol. 55, issue 5, pp. 1046-1057.
- Gli scienziati usano la paleontologia digitale per produrre la voce del dinosauro Parasaurolophus, dai Sandia National Laboratories (1997-12-05). Recuperato il 20 gennaio 2009.
- Diegert, Carl F. e Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen from the Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. e Williamson, Thomas E., 1996. Un nuovo cranio di Parasaurolophus (forma a cresta lunga) dal Nuovo Messico: caratteristiche esterne e interne (TAC) e loro implicazioni funzionali, dal Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborata struttura di raffreddamento del SNC nei grandi dinosauri Journal of Nature, vol. 275, alle pp. 441-443.
- Maryańska, Teresa e Osmólska, Halszka, nel 1979. Aspetti dell’anatomia cranica degli adrosauri, dal Journal of Lethaia, vol. 12, alle pp. 265-273.
- Gli scienziati usano la paleontologia digitale per produrre la voce del dinosauro Parasaurolophus; dai Sandia National Laboratories.
- Restauri di P. walkeri e P. cyrtocristatus, alla stessa scala, da Scott Hartman; su Skeletal Drawing.com.
- Pagina del Parasaurolophus su Enchanted Learning.com