
主に海洋動物のいくつかのグループは、最上級の言葉を使わずに議論するのは困難です。 最も偉大な」、「最も成功した」、「最も重要な」といった形容詞が、これらの動物についての議論を支配する傾向があります。 このような最上級の動物は、主として、大きい、目立つ、生態学的に成功した、あるいは特定の著者のお気に入りである、という傾向がある。 また、多くの動物が豊富であり、その活動によって私たちの注目を集める動物もいる。 このコラムで取り上げる両脚類は、多くの点で、最上級の表現を用いて論じられるこれらの動物群に当てはまるはずです。 両脚類は確かに「生態学的成功者」であり、また進化的にも非常に成功した動物である。 しかし、両生類は一般に小さく、不明瞭で、しばしば不可解な色をしている。 特に熱帯地方では、もっと美しい動物や印象的な動物がたくさんいるため、これらの「属性」が相まって、彼らの自然史はあまり知られていない。 しかし、このような地域でも、両脚類は多様で豊富であり、生態学的に重要です。 したがって、サンゴ礁の水槽に最もよく生息しているのが両脚類であることは明らかでしょう。 ほとんどの場合、両脚類は水槽に入れるのに最も望ましい動物であることもわかりました。
では、両脚類とは何なのでしょうか? さて、伝統によれば、「なぜアヒルは?」という古いイギリスの謎かけの答えは、「足の1本が両方とも同じだから」です。 同じように、”両脚類はなぜ?”の答えは、”脚の2本が両方とも違うから “であろう。 両脚類という名前は、”amph”(ギリシャ語のamphi=amphisで「両側の、二重の、また離れた、離れた、丸い」という意味)と “pod”(ギリシャ語のpousまたはpodosで「足」という意味、podion「小さな足」、podotēs「足を持つ」のように)からきており、これらの動物は表面的には2種類の付属器または足を持っているということ(Jaeger、1955)である。
分類学的には、両脚類は節足動物門甲殻綱上綱両脚目(Order Amphipoda)に属し、その分類学上の位置は次のとおりです。 無脊椎動物学者にとって、この用語の羅列は非常に多くの情報を与えてくれる。 しかし、多音節の単語のリストを読んだ後、平均的なリーフアクアリストは、アヒルについての古い英語の謎かけの方がより意味があると思うようになるのではないかと思います。 両脚類は節足動物であり、節足動物の特徴である、節がある体、節がある付属器、外骨格、つまり外皮を備えているのである。 表皮は、動物の外側の表皮と、その表皮から分泌される多くの化学物質で構成されている。 この外骨格は柔軟ではあるが伸縮自在ではないので、成長するためには頻繁に脱皮、つまり「皮」を脱いで新しい「皮」を生やすことが必要である。 実際には、皮膚そのものが剥がれるのではなく、古い外側の化学層であるクチクラが部分的に化学的に分解され、再吸収されるのである。 残ったもの、つまり元の皮膚の薄い残骸が剥がれ落ちるのです。 表皮はキューティクルの下にあり、動物の他の部分と一緒に残っている。 脱皮の過程で、古いクチクラの下に新しい、より大きなクチクラが形成され、残存する角質が剥がれ落ちて初めて露出する。 新しいキューティクルは最初は柔らかいが、しばらくすると硬くなる。
節足動物の主なグループである甲殻類(エビ、カニ、両脚)、単弓類(昆虫、ムカデ)、鋏角類(クモ、ダニ、カブトガニ)はそれぞれ非常に異なっており、多くの独自の特徴を示している。 これらの動物の祖先はおそらく三葉虫のようなものであるが、その祖先は約5億年前に生きており、それ以来、各系統の動物は多くの分岐と多くの改変が行われたのである。 両者を分ける違いの多くは明白である。 例えば、両生類を含む甲殻類には必ず2対の触角があるが、単弓類には1対しかなく、鋏角類には全くない。 また、甲殻類の付属器は一般に2つの部分、または枝から構成されている。 例えば、カニの歩脚は2本の枝からなり、1本目は脚の見える部分、2本目は脚の付け根から分岐している鰓(えら)である。 甲殻類以外の節足動物で枝分かれした付属物を持つものはいない。 甲殻類は主に海産動物であり、陸上でも淡水でもそこそこ成功しているグループである。 昆虫類と鋏角類は陸上や淡水域が主体である。 7255>
表面的にはカニやエビとかなり違うように見えるが、両脚類は両グループに比較的近いと考えられている。 そのため、両脚類は “近海エビ “と呼ばれるグループに分類されています。 これらの種類の動物はすべて、体の各部位に見られる付属肢の数が同じであることや、一般的な体の形など、多くの類似した構造的特徴を有している。 しかし、両生類やその他の小動物のグループには、カニやエビに見られるような特徴的な甲羅がない。 また、両生類にはメスの腹面に子袋があるが、カニやエビには子袋がないことも重要な違いである。 両脚類のメスは交尾の後、卵を子袋に入れ、両脚類として完全に機能するようになるまで卵の世話をする。 つまり、両脚類には自由生活する幼生がおらず、成体(主にメス)の実際の移動によって分散する必要があるのです。
両脚類には3つの大きなグループと1つの非常に小さなグループがありますが、私たちの水槽で普通に見られるのは、そのうちの1つのグループの代表だけでしょう。 一般的な両脚類は「ガンマー型」と呼ばれ、淡水でよく見られる両脚類の属名Gammarusにちなんで名づけられたグループに属します。 このような動物は、体の前部に甲羅がないことで特徴づけられ、エビと区別される。 また、体節が分かれているのも特徴である。 エビやカニでは、体節が甲羅に覆われていたり、融合していたりするのと対照的である。
外形
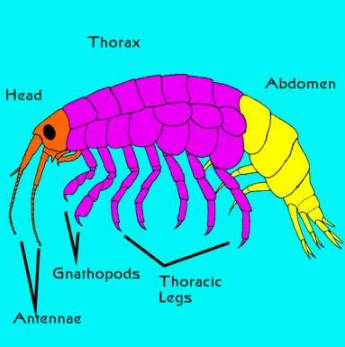
両脚類は一般に左右に扁平で、頭の両側に大きな複眼を持つ。 エビやカニの目とは異なり、この目は茎になく、頭の中に埋め込まれています。 さらに面白いことに、両生類という名前は2種類の脚を指しているが、実際には1つの動物に様々な脚の構造タイプがある。 両脚類という名前は2種類の脚を指しているが、実際には1つの動物に様々な脚の構造がある。 胸部の最も前方にある2つの節には、ニワトポッドと呼ばれる付属肢があり、これが変化している。 「Gnathは「顎」を意味し、顎脚は文字通り「顎の足」で、摂食に使われると考えられているが、このグループの動物について詳しく注意深く観察されたことは比較的少ない。 しかし、このグループの動物をじっくり観察したことはほとんどない。この付属肢が実際に何に使われているかは、ほとんど疑問の余地がある。 いずれにせよ、顎脚の先端には「亜門」と呼ばれるジャックナイフのような反り返った爪があるのが普通である(図2、6、7参照)。 2対の顎脚の後方には2対の短い脚があり、その後方には3対の長い脚がある。 長い方の脚は外側に広がる傾向があり、自転車の車輪のような典型的な姿勢であることがわかる。
 |
 |
|
図2. 一般的な両生類の外形解剖図。 頭部は赤、胸部は紫、腹部は黄色で示す。
|
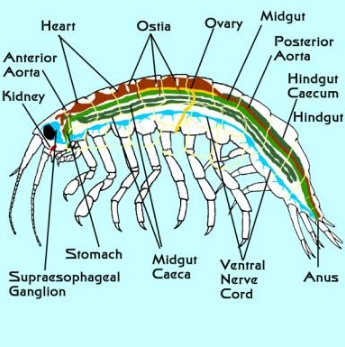
図3. 一般的な両生類(Gammaridean)の内部解剖。 心臓は茶色、腸の各部は緑、神経系は青、生殖腺と生殖管は黄色、腎臓(腎腺)はピンクである。
|
内部解剖
典型的な両生類の内部解剖は図3に示すとおりである。 すべての節足動物のように、神経系は腹面の中央部に沿ってある。 各節に神経節腫脹がある。 大きな食道上神経節は食道の上にあり、食道の周囲を走る神経とともに脳を構成している。 目は大きな視神経でこれらの神経節と直接連絡している。 触角は感覚器であり、ここにも大きな神経が走っている。
口は頭の付け根近くにあり、短い食道は頭のすぐ後ろにある胃に垂直に通じている。 胃の中にはキチン質の板がいくつか並んでいて、食物をすりつぶす役目をしている。 外骨格が硬いため口の大きさが制限され、入ってきた食べ物のほとんどは液体か、口の周りの付属器官で細かく引き裂かれる。 長い中腸は体の大部分を貫いている。 中腸から一連の袋(カイコ)が生じている。 非常に短い1本の盲腸が腸の上部から立ち上がり、頭部に向かって短い距離を走る。 2〜8個の袋が胃のすぐ後ろの中腸の側面と底面から生じている。 これらは後方を通って、ほぼ腹部まで伸びている。 消化と消化酵素および “ジュース “の分泌は、さまざまな袋の中で行われる。 中腸の末端には同じような袋が1つあり、中腸の小嚢と生殖腺の上を前方に向かって伸びている。 その機能は不明である。 この盲腸の起始部より後方では、腸は後腸と呼ばれる。
胸部には長い管状の心臓が背側の体壁から吊り下げられている。 3対の「オスティア」と呼ばれる弁のついた開口部によって、血液は周囲の空間から心臓に一方的に流れ込んでいる。 心臓が収縮すると、血液は前大動脈または後大動脈からそれぞれ前方および後方に送り出される。 側副血行路は心臓から体壁に通じている。 血液はこれらの血管を出て、チャネルで組織の周りや中を流れます。 血流は速く、小さな両生類ではわずか数秒で1周してしまう。 血液にはさまざまな種類の角質が含まれているが、そのほとんどがどのように機能しているのか、実はわかっていない。 両生類は特定の呼吸器官を持たず、ガス交換はおそらく体表全体で行われている。
両生類はかなり単純なライフサイクルを持っているため、我々のシステムでうまく繁殖することができる。 一般に雌雄が分かれており、両性具有はまれである。 雌雄の区別は容易である。 卵管は第6胸節の脚の付け根に、精管は第8胸節の脚の付け根に開口している。 雄は一対のペニスを持ち,胸部付属器が変化し,受精は体内で行われる。 眼、触角、第2鰓脚の形状も雌雄で異なる場合がある。 また、メスには先に述べた子袋がある。 この袋は、メスの胸脚の内側の縁を起点とする一連の板で、メスの底面の下に形成されている。 この板は腹部正中線を越えて伸び、反対側の板と重なり、腹面との間に部屋を作る。 交尾の後、受精卵はこの部屋に預けられ、そこで保持されながら成長する。 幼虫が成熟して稚魚になると、メスは子ガメを放し、子ガメはその周辺に分散していく。 両脚類には、自由遊泳や自由生活する幼生期がない。 このような生殖では、もちろんメスが食べられない限り、幼虫が死亡する可能性はあまりない。 通常、個体数が増えて密になると、妊娠したメスが移住して新たな個体群を作る。
認識
一般に、両生類は上の図2の図や下の写真(図4)のようにかなり似ていると見なされている。 しかし、これは大きなグループで、おそらく5,000種を優に超える種があり、中には奇妙で壮大な変わり種もあります。 また、形や大きさもかなりバラエティに富んでいる(図5)。 もちろん、これらの動物が多様な生態的ニッチを占めていることは言うまでもない。 両生類は、捕食者、草食者、腐食者、細菌食者である場合がある。 雑食性であるものもある。 多くは他の動物に寄生し、寄生虫もいる。
 |
 |
のもので、両脚類の特徴的な体型をしている。

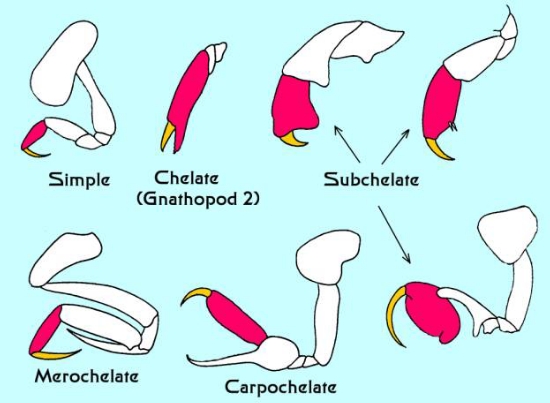
どんな動物でも、その自然史の中で最も重要なのは食物を得る必要性であり、両生類の摂食方法の多様性は、基本的な摂食付属物の違いに反映されていると思われる。 一般に、両脚類は獲物の捕獲、餌の操作、あるいは摂食に関与していると考えられている。 顎脚の基本的な形態は図2、図7の図と写真、図6に例示されているが、実際の構造がどうなっているかはかなりのバリエーションがある。 図7に顎脚の様々な形状の一部を示す。 図2に示した単純な亜門の付属器からの変化は、それぞれ食生活や生活習慣の違いを反映している。 その結果、両脚類は多くの種類に分化し、水族館では変わった種類に出会うこともある。 両生類の亜門型第1節足動物。 この付属肢は透明で、
前脚に対して乳頭を閉じるための比較的大きな筋肉が
表示され、血流路の中の血球も表示されている。
非羊膜類両脚類
奇妙な両脚類の非常に小さなグループにIngolfiellideaと呼ばれるものがあります。 これらの動物は通常、砂粒の間や上に住んでいて、私の知る限り、水族館では一度も見たことがありません。 他の2つのグループは、生態学的に重要で多様な両脚類です。 それらはHyperiideaとCaprellideaです。 両脚類はプランクトンの中に生息し、多くはゼラチン質の動物性プランクトンを捕食または寄生しています。 ハイペリアイデスの中にはかなり奇妙なものもあり、フロニマという生物は、同名の映画に出てくるエイリアンのモデルになったという噂がある。 Phronimaは遠洋性で、遠洋性の鮪の家に住んでいる。
両脚類の他のグループであるCaprellidsは、魅力的な動物である。 カプレロイドの1つの系統はクジラに住み、”クジラシラミ “と呼ばれている。 もう一つの系統はスケルトンシュリンプと呼ばれ、これは適切な名前である。 小さな多腕の骸骨のような姿をしており、藻類やウミウシの背中、ゴルゴニアンに付着しているのを見かける。
ガンマー類を典型的な両生類と考えることに慣れている人にとって、初めてカペルリッドを調べたとき、これを両生類と考えるのは非常に難しいことである。 一見して、そしておそらく二度目、三度目にも、カプレロイドはガンマー類とは全く違うように見えるのです。 長い管状の体は小さいが球根状の頭部で終わり、2対の長い触角が生えており、その長さは体の他の部分と同じくらいかもしれない。 棒のように見えるが、ジャックナイフのように閉じる爪で終わる、比較的大きく顕著な付属器官を持っている。 スケルトンシュリンプの名前は適切である。 ある種の奇妙な甲殻類の骨格が生き返ったように見えるのです。
大きいものでは数センチになることもあるが、リーフタンクに入っているものはほとんどが1センチ以下のオーダーである。 前脚を伸ばすと体高と同じぐらいの幅になることが多い。 岩場や藻の上に座り、後脚でつかむ習性がある。 体は水中に垂直に伸び、前肢を大きく広げて、水中で何かが通り過ぎるのを待つ。 他の両生類と同様に、メスには胞子嚢がある。 この場合、動物が立ち上がったときに体の真ん中にあり、いくつかの動物の真ん中に小さな白い点として見えることが多い。 一般的には淡い琥珀色や白色であるが、他の色を示すこともある。 これらの多くは無害または有益な草食動物またはスカベンジャーであるが、一部は肉食性であり、小さなソフトコーラルのポリプやその他の小動物を食べることができる。
多くの動物と同様に、カプレロイドはライブロック、藻類、サンゴ、ライブサンドの上でヒッチハイカーとして私たちのシステムに入ることがあります。 一般的に、彼らはかなり無害であり、非常に多くの魚の餌となっています。 時折、小さな問題を引き起こすものも発見される。 草食性のカプレロイドは、適切な餌(一般的には藻類)があれば、レフュジアムやサンプ、時にはメインタンクで培養することができる。 培養は光と藻類を供給し、動物にその行動をさせる。 肉食の個体は、問題を起こすようであれば、鉗子で取り除くことができる。 一般に、メイン水槽では、魚がその存在に気づかないうちに取り除いてしまう。 カペル型両生類。 左:オス。 右:雌(大きな袋の中に稚魚がいることに注意)。
水槽での行動とケア
水槽にいる両生類の大部分を見分けるのは一般的にとても簡単です。 また、胸部後方の付属肢が横方向に伸びているのも特徴的である。 水族館で見られる種は小型で、体長数mm程度になることはほとんどない。 通常、図2のイラストとほとんど変わらない。
これらの虫は単にガンマーシュリンプと呼ばれることも多いが、これは誤用で、水槽で見られる種のほとんどはガンマーシュリンプの仲間ではない。 また、「グランパス」や「グラマス」など、「ガンマー」の語源となる言葉もよく耳にします。 ほとんどのアクアリストは、すべての両生類は水槽で見る標準的なガンマー系両生類とほぼ同じで、これらの動物はすべて草食性か腐食性であると考えています。 リーフタンクという限られた世界の中では、これは多かれ少なかれ正しいのですが、すべてのガンマー系両脚類が草食性というわけではなく、中には非常にはっきりとした肉食性のものもあります。 植物や藻類を好んで食べる傾向があり、藻類をかすめるか、植物や藻類由来の残骸を食べます。 一般的に、動物の肉はあまり食べませんが、時折、捕食性のある両脚類が体内に入ることがあります。 これらの種は顕微鏡で観察しないと区別がつきにくいので、ほとんどの趣味家はこの2つの種を区別する唯一の方法は(それぞれの種に数百の可能性があることに留意してください)、餌を食べるところを観察することなのです。 私たちのアクアリウムでは、端脚類は通常清掃係の一員です。 さらに、両脚類はどんな魚でも捕まえることができる良い餌です。 7255>
Man-Eating Amphipods and Other Oddities
 |
 |
 |
両脚類の肉食性は珍しいことではなく、時には本当に肉食性のものが海の水族館にヒッチハイクでやってくることもあります。 深海では、あるいは光帯以下の多くの浅い水域でも、肉食性の両脚類は清掃ギルドの主要メンバーであるか、それ自体で捕食しています。 私は、時には人を食べる種さえ知っています。 この両脚類の一種、Chromopleustes pugettensisの色彩は印象的である(図10)。 体色は濃厚な暗褐色で、鮮やかな白色の鞍部と金色の縦縞がある。 目はラベンダー色で、脚は青色である。 このコラムの常連なら、この色彩を警告色あるいはアポセマティック色彩と認識することだろう。 この色彩は危険な動物の色彩であり、この両生類は捕食者にとっても獲物にとっても明らかに危険である。 さらに、この両脚類は隠れない種である。 昼間でも底にいることがよくわかるし、近づいても逃げない。 この行動も捕食から守られている証拠である。

Chromopleustes pugettensisは分布が斑点状である。 一般的には珍しいが、ある地域で発見された場合、大きな群れで発見される傾向があり、私は1回のダイビングで20,000個体以上の群れを観察したことがある。 一年の大半、Chromopleustes pugettensisはナマコと一緒にいるようである。 詳しい生態は不明だが、小さなナマコや大きなナマコの一部を食べているようである。 ナマコは多くの棘皮動物と同様、サポニンと呼ばれる有毒な化学物質を含んでいる。 サポニンは多くの棘皮動物に捕食の自由を与えると考えられており、一般に棘皮動物を捕食するものは非常に稀である。 このため、クロモプルスティスはナマコを食べることに適応し、ナマコのサポニンを体内に濃縮しているのではないかと推測されている。 しかし、一連の実験では、この地域のほとんどの魚がこの両生類を食べないことが明らかになった。 何年も前に、私はこの端脚類をさまざまな捕食者に食べさせようとする実験をしたことがあります。 魚がその虫を食べたのは一度だけで、すぐに吐き戻されました。 翌朝、その魚は死んでいた。 7255>
1983年4月2日、私はワシントン州のサンファン諸島にあるポール・パスという地域でダイビングをしていました。 このダイビングの間、パートナーと私は大きなウミウシ、Pycnopodia helianthoidesが産卵しているところに出くわしました。 そのウミウシは端脚類の大群に完全に覆われており、上面の一部を引きちぎっているような状態だった。 近寄ろうとすると、群れが盛り上がってきて、その一部が私の顔にかかり、何が起こったのかわからないうちに、虫が私の顔や唇を噛んでいるのである。 私は急いで「バックペダル」をして、なんとか虫を払い落としましたが、その間に何箇所か皮膚が破れてしまい、大量に出血してしまいました。 NASTY LITTLE BUGS!!!
幸いなことに、ほとんどのリーフアクアリウムの両脚類はもっと良性です。 しかし、上記の経験から、ほとんどのアクアリストは、両脚類が捕食しないことを当然のことと思ってはいけないと思うはずです。 一方、引っ込み思案で警告色を持たない両生類は、おそらく安全で私たちのシステムにとって有益な存在です。