

Parasaurolophus (pronuncia-se /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, normalmente também /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; que significa “quase lagarto cristalizado” em referência ao Saurolophus) é um gênero de dinossauro ornitopod do Período Cretáceo Final do que é hoje a América do Norte, há cerca de 76-73 milhões de anos. Era um herbívoro que caminhava tanto como bípede como quadruplicado. Três espécies são reconhecidas: P. walkeri (a espécie do tipo), P. tubicen, e o P. cyrtocristatus de crista curta. Os remanescentes são conhecidos de Alberta, Canadá, e do Novo México e Utah, EUA. Foi descrito pela primeira vez em 1922 por William Parks de um crânio e esqueleto parcial em Alberta.
Parasaurolophus é um hadrosaurido, parte de uma família diversa de dinossauros Cretáceos conhecidos pela sua gama de ornamentos de cabeça bizarros. Este género é conhecido pela sua grande e elaborada crista craniana, que na sua maior forma forma um longo tubo curvo projectando-se para cima e para trás a partir do crânio. O Charonosaurus da China, que pode ter sido o seu parente mais próximo, tinha um crânio semelhante e potencialmente uma crista semelhante. A crista tem sido muito discutida pelos cientistas; o consenso é que as principais funções incluíam o reconhecimento visual de ambas as espécies e sexo, ressonância acústica e termorregulação. É uma das mais raras, sendo conhecida apenas de um punhado de bons exemplares.
Parasaurolophus está entre os dinossauros mais reconhecidos, sendo frequentemente retratada em programas de televisão, filmes, livros, mercadorias e outros meios de comunicação relacionados aos dinossauros. Alice Perry
Descrição

O tamanho do Parasaurolophus walkeri comparado a um humano.
Como é o caso da maioria dos dinossauros, o esqueleto do Parasaurolophus é incompletamente conhecido. O comprimento do espécime do tipo de P. walkeri é estimado em 9,5 metros (31 pés). Seu crânio tem cerca de 1,6 metros de comprimento, incluindo a crista, enquanto o crânio do tipo de P. tubicen tem mais de 2,0 metros de comprimento, indicando um animal maior. O seu peso é estimado em 2,5 toneladas (2,7 toneladas). Seu único membro anterior conhecido era relativamente curto para um hadrosaurido, com uma omoplata curta mas larga. O fémur mede 103 centímetros de comprimento em P. walkeri e é robusto pelo seu comprimento quando comparado com outros hadrosaurídeos. O antebraço e os ossos pélvicos também foram fortemente construídos.
Como outros hadrosaurídeos, era capaz de andar em duas ou quatro pernas. Provavelmente preferiu forragear em quatro patas, mas correu em duas. As espinhas neurais das vértebras eram altas, como era comum nos lambeosauros; as mais altas sobre os quadris, aumentavam a altura das costas. As impressões cutâneas são conhecidas por P. walkeri, mostrando escalas uniformes em forma de tubérculo mas sem estruturas maiores.
Parasaurolophus walkeri com detalhe de escalada.
A característica mais notável foi a crista craniana, que se projetava da parte posterior da cabeça e era composta pela pré-maxila e ossos nasais. O espécime do tipo P. walkeri tem um entalhe nas espinhas neurais perto de onde a crista atingiria a parte posterior, mas esta pode ser uma patologia peculiar a este indivíduo. William Parks, que deu o nome ao gênero, levantou a hipótese de que um ligamento corria da crista até o entalhe para apoiar a cabeça. Embora a idéia pareça improvável, Parasaurolophus às vezes é restaurado com uma aba de pele da crista até o pescoço.
A crista era oca, com tubos distintos que levavam de cada narina até o final da crista antes de inverter a direção e descer a crista de volta para dentro do crânio. Os tubos eram mais simples em P. walkeri, e mais complexos em P. tubicen, onde alguns tubos eram cegos e outros se encontravam e se separavam. Enquanto P. walkeri e P. tubicen tinham cristas longas com apenas uma ligeira curvatura, P. cyrtocristatus tinha uma crista curta com um perfil mais circular.
Classificação
Como o seu nome indica, inicialmente pensou-se que Parasaurolophus estava intimamente relacionado com Saurolophus por causa da sua crista superficialmente semelhante. Entretanto, logo foi reavaliado como membro da subfamília lambeosaurina de hadrosaurídeos – Saurolophus é uma hadrosaurina. É geralmente interpretado como um ramo separado dos lambeosaurinos, distinto do Corythosaurus, Hypacrosaurus e Lambeosaurus. Seu parente mais próximo conhecido parece ser o Charonossauro, um lambeossauro com um crânio semelhante (mas ainda sem crista completa) da região de Amur no nordeste da China, e os dois podem formar um clade Parasaurolophini. P. cyrtocristatus, com sua crista curta e mais arredondada, pode ser a mais basal das três espécies de Parasaurolophus conhecidas, ou pode representar espécimes subadultos ou fêmeas de P. tubicen.
História
Dicovery and naming
Cast do espécime incompleto do tipo Parasaurolophus walkeri em Warszawa.
Meaning “near cristted lizard”, o nome Parasaurolophus deriva do grego para/παρα “ao lado” ou “perto”, saurus/σαυρος “lizard” e lophos/λοφος “crest”. É baseado na ROM 768, um crânio e esqueleto parcial sem a maior parte da cauda e as pernas traseiras abaixo dos joelhos, que foi encontrado por uma festa de campo da Universidade de Toronto em 1920 perto de Sand Creek ao longo do Rio Red Deer em Alberta, Canadá. Estas rochas são agora conhecidas como o Parque de Formação de Dinossauros do Alto Cretáceo da Era Campaniana. William Parks nomeou o espécime P. walkeri em homenagem a Sir Byron Edmund Walker, Presidente do Conselho de Curadores do Museu Real Ontário. Restos de Parasaurolophus são raros em Alberta, com apenas um outro crânio parcial (provavelmente) da Formação do Parque dos Dinossauros, e três espécimes do Parque dos Dinossauros sem caveiras, possivelmente pertencentes ao gênero. Em algumas listas faunísticas, há uma menção de possível material P. walkeri no Hell Creek Formation of Montana, uma unidade rochosa do final da era Maastrichtian. Esta ocorrência não é notada por Sullivan e Williamson em sua revisão de 1999 do gênero, e não foi mais elaborada em outros lugares.
Em 1921, Charles H. Sternberg recuperou um crânio parcial (PMU.R1250) do que é agora conhecido como a Formação Kirtland ligeiramente mais jovem no Condado de San Juan, Novo México. Este espécime foi enviado para Uppsala, Suécia, onde Carl Wiman o descreveu como uma segunda espécie, P. tubicen, em 1931. O epíteto específico é derivado do latim tǔbǐcěn “trompetista”. Um segundo, quase completo crânio de P. tubicen (NMMNH P-25100) foi encontrado no Novo México, em 1995. Usando tomografia computorizada deste crânio, Robert Sullivan e Thomas Williamson deram ao gênero um tratamento monográfico em 1999, cobrindo aspectos de sua anatomia e taxonomia, e as funções de sua crista. Williamson publicou posteriormente uma revisão independente dos restos mortais, discordando das conclusões taxonômicas.
John Ostrom descreveu outro bom espécime (FMNH P27393) do Novo México como P. cyrtocristatus em 1961. Ele inclui um crânio parcial com uma crista curta e arredondada, e grande parte do esqueleto pós-craniano, exceto os pés, pescoço e partes da cauda. Seu nome específico é derivado do latim curtus “encurtado” e cristatus “cristatus cristalizado”. O espécime foi encontrado ou no topo da Formação de Fruitland ou, mais provavelmente, na base da Formação de Kirtland sobrevoando. A variedade desta espécie foi expandida em 1979, quando David B. Weishampel e James A. Jensen descreveram um crânio parcial com uma crista semelhante (BYU 2467) da Formação Kaiparowits da era Campaniana do Condado de Garfield, Utah. Desde então, outro crânio foi encontrado em Utah com a morfologia da crista curta/redonda de P. cyrtocristatus.
Espécie
Parasaurolophus walkeri skull cast, Museu de História Natural, Londres.
Parasaurolophus cyrtocristatus skull cast, Field Museum of Natural History.
O tipo de espécie P. walkeri, de Alberta, é conhecido a partir de um único espécime. Difere de P. tubicen por ter tubos mais simples na sua crista, e de P. cyrtocristatus por ter uma crista longa e não arredondada e um braço mais longo do que o antebraço.
P. tubicen, do Novo México, é conhecido pelos restos de pelo menos três indivíduos. É a maior espécie, com passagens de ar mais complexas na sua crista do que P. walkeri, e uma crista mais longa e reta do que P. cyrtocristatus.
P. cyrtocristatus, do Novo México e Utah, é conhecida a partir de três exemplares possíveis. É a espécie mais pequena, com uma crista curta e arredondada. O seu pequeno tamanho e a forma da sua crista levaram vários cientistas a sugerir que representa juvenis ou fêmeas de P. tubicen, que é aproximadamente da mesma época e da mesma formação no Novo México. Como foi observado por Thomas Williamson, o tipo de material de P. cyrtocristatus é cerca de 72% do tamanho de P. tubicen, próximo do tamanho em que outros lambeosaurines são interpretados para começar a mostrar dimorfismo sexual definitivo em suas cristas (~70% do tamanho adulto). Esta posição foi rejeitada em revisões recentes de lambeosaurines.
Paleobiologia
Paleoecologia
Gorgosaurus (esquerda) atacando um Parasaurolophus cyrtocristatus.
Parasaurolophus walkeri, da Formação Parque dos Dinossauros, era membro de uma fauna diversificada e bem documentada de animais pré-históricos, incluindo dinossauros bem conhecidos como o Centrossauro com chifres, Estracosaurus, e Chasmosaurus; os companheiros Prosaurolophus, Gryposaurus, Corythosaurus e Lambeosaurus; o Gorgosaurus tiranossauro; e o Edmontonia e Euoplocephalus blindados. Era um constituinte raro desta fauna. A Formação do Parque dos Dinossauros é interpretada como um cenário de baixo relevo de rios e planícies aluviais que se tornaram mais pantanosos e influenciados pelas condições marinhas ao longo do tempo, à medida que o mar interior ocidental transgredia para oeste. O clima era mais quente que o actual Alberta, sem geadas, mas com estações mais húmidas e secas. As coníferas eram aparentemente as plantas dominantes da copa das árvores, com um subcoberto de fetos, samambaias e angiospermas.
As novas espécies mexicanas compartilharam seu ambiente com os grandes saurópodes Alamosaurus, Kritosaurus de pato, Pentaceratops de chifres, Nodocephalosaurus blindados, Saurornitholestes, e atualmente não denominados tiranossauros. A Formação de Kirtland é interpretada como planícies de inundação de rios que aparecem após um recuo do mar interior ocidental. Coníferas eram as plantas dominantes, e dinossauros chasmosaurinos eram aparentemente mais comuns que os hadrosaurídeos.
Alimentação
Como um hadrosaurídeo, Parasaurolophus era um grande herbívoro bípede/quadrupedal, comendo plantas com um crânio sofisticado que permitia um movimento de moagem análogo à mastigação. Seus dentes eram continuamente substituídos e embalados em baterias odontológicas que continham centenas de dentes, dos quais apenas um punhado relativo estava em uso a qualquer momento. Utilizava o seu bico para cultivar material vegetal, que era segurado nos maxilares por um órgão semelhante a uma bochecha. A alimentação teria sido feita a partir do solo até cerca de 4 metros acima. Como observado por Bob Bakker, os lambeosaurines têm bicos mais estreitos que os hadrosaurines, implicando que Parasaurolophus e seus parentes poderiam alimentar-se de forma mais seletiva que seus parentes de bico largo e sem cristas.
Crista cranial
Muitas hipóteses foram avançadas quanto às funções que a crista cranial do Parasaurolophus desempenhava, mas a maioria foi desacreditada. Acredita-se agora que ela possa ter tido várias funções: exibição visual para identificação de espécies e sexo, amplificação sonora para comunicação, e termorregulação. Não está claro qual foi o momento mais significativo na evolução da crista e suas passagens nasais internas.
Diferenças entre espécies e estágios de crescimento
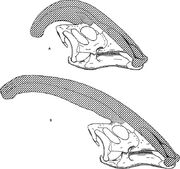
Diagrama comparando as cristas nariais de Parasaurolophus cyrtocristatus (a) e Parasaurolophus walkeri (b).
Como para outros lambeosaurines, acredita-se que a crista craniana de Parasaurolophus mudou com a idade e foi uma característica sexualmente dimórfica em adultos. James Hopson, um dos primeiros pesquisadores a descrever cristas de lambeosaurinos em termos de tais distinções, sugeriu que P. cyrtocristatus, com sua pequena crista, era a forma feminina de P. tubicen. Thomas Williamson sugeriu que era a forma juvenil. Nenhuma das hipóteses se tornou amplamente aceita. Como apenas seis bons crânios e uma braquicase juvenil são conhecidos, material adicional ajudará a esclarecer essas potenciais relações. Williamson observou que, de qualquer forma, o Parasaurolophus juvenil provavelmente tinha cristas pequenas e arredondadas como P. cyrtocristatus, que provavelmente cresceram mais rapidamente à medida que os indivíduos se aproximavam da maturidade. O recente reestudo de uma braincase juvenil anteriormente atribuída a Lambeosaurus, e agora atribuída a Parasaurolophus, fornece evidências de que uma pequena crista tubular estava presente nos juvenis. Este espécime preserva uma pequena queima ascendente dos ossos frontais que era semelhante mas menor do que a observada nos espécimes adultos; nos adultos, os frontais formavam uma plataforma que suportava a base da crista. Este espécime também indica que o crescimento da crista em Parasaurolophus e o perfil facial dos indivíduos jovens diferem do modelo do Corythosaurus-Hypacrosaurus-Lambeosaurus, em parte porque a crista de Parasaurolophus carece da fina “coxcomb” óssea que compõe a porção superior da crista dos outros três lambeosaurinos.
Hipóteses rejeitadas sobre a função
Muitas sugestões iniciais focalizadas em adaptações para um estilo de vida aquático, seguindo a hipótese de que os hadrosaurídeos eram anfíbios, uma linha de pensamento comum até os anos 60. Assim, Alfred Sherwood Romer propôs que servisse como um snorkel, Martin Wilfarth que fosse um acessório para uma probóscide móvel usada como tubo respiratório ou para coleta de alimentos, Charles M. Sternberg que servisse como um filtro de ar para manter a água fora dos pulmões, e Ned Colbert que servisse como reservatório de ar para estadias prolongadas debaixo d’água.
Outras propostas eram de natureza mais física. Como mencionado acima, William Parks sugeriu que ela fosse unida às vértebras com ligamentos ou músculos, e que ajudasse a mover e apoiar a cabeça. Othenio Abel propôs que ela fosse usada como arma em combate entre membros da mesma espécie, e Andrew Milner sugeriu que ela poderia ser usada como um defletor de folhagem, como a crista do capacete (chamado “casque”) da batina. Ainda outras propostas fizeram do alojamento de órgãos especializados a principal função. Halszka Osmólska sugeriu que abrigou glândulas salinas, e John Ostrom sugeriu que abrigou áreas expandidas para o tecido olfativo e muito melhorou o olfato dos lambeosaurines, que não tinham capacidades defensivas óbvias. Uma sugestão incomum, feita pelo criacionista Duane Gish, é que as glândulas químicas da crista alojava jatos de “fogo” químico em inimigos, semelhante ao escaravelho bombardeiro dos tempos modernos.
A maior parte destas hipóteses foram desacreditadas ou rejeitadas. Por exemplo, não há nenhum buraco no final da crista para uma função de snorkeling. Não há cicatrizes musculares para uma probóscide e é duvidoso que um animal com um bico precise de uma. Como uma câmara de ar proposta, ele não teria mantido fora de água. O reservatório de ar proposto teria sido insuficiente para um animal do tamanho de Parasaurolophus. Outros hadrosaurídeos tinham cabeças grandes sem precisar de grandes cristas ocas para servir como pontos de fixação para os ligamentos de suporte. Além disso, nenhuma das propostas explica por que a crista tem tal forma, por que outros lambeossauros deveriam ter cristas que parecem muito diferentes mas desempenham uma função similar, como os hadrossauros sem cristas ou com cristas sólidas se davam sem tais capacidades, ou por que alguns hadrossauros tinham cristas sólidas. Estas considerações impactam particularmente as hipóteses baseadas no aumento das capacidades dos sistemas já presentes no animal, tais como a glândula salina e as hipóteses de olfacto, e indicam que estas não eram funções primárias da crista. Além disso, trabalhos sobre a cavidade nasal dos lambeosauros mostram que os nervos olfativos e o tecido sensorial correspondente estavam em grande parte fora da porção das passagens nasais da crista, portanto a expansão da crista teve pouco a ver com o olfato.
Funções sociais
Restauração da vida de Parasaurolophus walkeri com hipotéticos folhos e faixas na crista.
Em vez disso, as funções sociais e fisiológicas tornaram-se mais apoiadas como função(ões) da crista, concentrando-se na identificação visual e auditiva e na comunicação. Como um objeto grande, a crista tem claro valor como um sinal visual, e diferencia este animal dos seus contemporâneos. O grande tamanho das tomadas oculares hadrosaurídeas e a presença de anéis escleróticos nos olhos implicam uma visão aguda e hábitos diurnos, evidência de que a visão era importante para estes animais. Se, como é comumente ilustrado, um folho de pele estendido da crista até o pescoço ou costas, a exposição visual proposta teria sido muito mais exposta. Como é sugerido por outros crânios de lambeosauros, a crista do Parasaurolophus provavelmente permitiu tanto a identificação das espécies (como separá-lo do Corythosaurus ou Lambeosaurus) quanto a determinação entre machos e fêmeas, com base na forma e tamanho.
Função de som
Modelo de computador das passagens nasais na crista do P. tubicen.
No entanto, a aparência externa da crista não corresponde à complexa anatomia interna das passagens nasais, o que sugere outra função contabilizada para o uso do espaço interno. Carl Wiman foi o primeiro a propor, em 1931, que as passagens serviam para uma função de sinalização auditiva, como uma crumhorn; Hopson e David B. Weishampel revisitaram esta idéia nos anos 70 e 80. Hopson descobriu que há evidências anatômicas que os hadrosaurídeos tinham uma audição forte. Há pelo menos um exemplo, no Corythosaurus relacionado, de um estribo esbelto (osso do ouvido reptiliano) no lugar, que combinado com um grande espaço para um tímpano implica em um ouvido médio sensível. Além disso, a lagena hadrosaurídea é alongada como a de um crocodiliano, indicando que a porção auditiva do ouvido interno estava bem desenvolvida. Weishampel sugeriu que P. walkeri foi capaz de produzir frequências de 48 a 240 Hz, e P. cyrtocristatus (interpretado como uma forma de crista juvenil) 75 a 375 Hz. Baseado na semelhança dos ouvidos internos dos hadrosaurídeos com os dos crocodilos, ele também propôs que os hadrosaurídeos adultos eram sensíveis a frequências altas, tais como a sua descendência poderia produzir. Segundo Weishampel, isto é consistente com pais e descendentes comunicando.
Modelagem computacional de um espécime bem preservado de P. tubicen, com passagens de ar mais complexas que as de P. walkeri, permitiu a reconstrução do possível som produzido na sua crista. O caminho principal ressoa em torno de 30 Hz, mas a complicada anatomia sinusal causa picos e vales no som.
Função de resfriamento
A grande superfície e vascularização da crista também sugere uma função termorreguladora. P.E. Wheeler sugeriu este uso pela primeira vez em 1978 como uma forma de manter o cérebro fresco. Teresa Maryańska e Osmólska também propuseram a termorregulação aproximadamente na mesma época, e Sullivan e Williamson se interessaram ainda mais. A discussão de David Evans sobre as funções da crista do lambeosaurine em 2006 foi favorável à idéia, pelo menos como um fator inicial para a evolução da expansão da crista.
Na Mídia
Parasaurolophus é um dinossauro muito popular. Foi no filme O Dinossauro da Disney como um membro do rebanho. Também fez várias aparições no famoso documentário Clash of the Dinosaurs. Também apareceu no popular programa de TV Parque Pré-histórico, onde se tornou presa de grandes carnívoros Deinosuchus e Albertosaurus. Apareceu nos filmes do Parque Jurássico, como membro do rebanho no primeiro filme, depois foi mantido em cativeiro pelos caçadores no segundo e correndo junto com o Corythosaurus no terceiro. e o personagem Dweeb em “We’re Back! A Dinosaur’s Story”, é o próprio Parasaurolophus.
Parasaurolophus também aparece em Turok, como um comedor dócil de plantas que normalmente não é prejudicial, mas pode ser agressivo se for severamente provocado.
Existe também um Parasaurolophus zord em Power Rangers: Dino Thunder e Power Rangers: Dino Charge.
Parasaurolophus aparece brevemente no início do filme da Disney Pixar The Good Dinosaur.
Veja também
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, página 229. Publicado: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Capítulo: Parasaurolophus, páginas 678-684. Publicado: 1997, McFarland & Co, em Jefferson, Carolina do Norte. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull and Wright, Hadrosaurian Dinosaurs of North America, pp. 209-213.
- Brett-Surman, Michael K. e Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks: Dinossauros Ceratopsianos e Ornitopsianos, capítulo: Anatomia apendicular em Campanian e Maastrichtian North American hadrosaurids, páginas 135-169. Publicado, 2006, Indiana University Press, em Bloomington e Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; e Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; e Dodson, Peter (eds.) The Dinosauria, 2ª edição, capítulo: Hadrosauridae, páginas 438-463. Publicado: 2004, University of California Press, em Berkeley. ISBN 0-520-24209-2
- 6,0 6,1 6,2 6,3 6,4 6,5 Parques, William A. Parasaurolophus walkeri, um novo gênero e espécie de dinossauro de trachodont cristalizado, volume 13, páginas 1-32. Publicado: 1922, University of Toronto Studies, Geology Series.
- 7,0 7,1 7,2 7,3 7,4 7,5 7,6 7,7 Sullivan, Robert M. e Williamson, Thomas E. Um novo crânio de Parasaurolophus (Dinosauria: Hadrosauridae) da Kirtland Formation of New Mexico e uma revisão do gênero, da série New Mexico Museum of Natural History and Science Bulletin, 15, páginas 1-52. Publicado: 1999, Museu de História Natural e Ciência do Novo México, em Albuqueque, Novo México.
- 8.0 8.1 8.2 Ostrom, John H. 1961 Uma nova espécie de dinossauro hadrosauriano do Cretáceo do Novo México, Journal of Paleontology, Volume 35, 3ª edição, nas páginas 575-577.
- Gilmore, Charles W., Sobre o gênero Stephanosaurus, com uma descrição do tipo de espécime do Lambeosaurus lambei, volume 38, edição 43, páginas 29-48, Parks. Publicado: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. e Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; e Dodson, Peter (eds.) The Dinosauria, 1ª edição, capítulo: Hadrosauridae, páginas 534-561. Publicado: 1990, University of California Press, Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., e Reisz, Robert R. 2007. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaurus (Ornithischia) from the Dinosaur Park Formation, Alberta, from the Journal of Vertebrate Paleontology, vol. 27, número 2, nas páginas 373-393.
- Godefroit, Pascal, Shuqin Zan; e Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., um dinossauro lambeosaurino do Maastrichtian tardio do nordeste da China, do Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, páginas 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Revisão de Hadrosauridae (Dinosauria, Ornithischia) da Bacia de San Juan, Novo México Lucas, S.G.; e Heckert, A.B. (eds.) Dinosaurs of New Mexico, do New Mexico Museum of Natural History and Science Bulletin, 17 Publicado pelo New Mexico Museum of Natural History and Science, em Albuquequeque, Novo México. Páginas 191-213.
- Liddell, Henry George e Robert Scott, 1980. Um Léxico Grego-Inglês (Edição abreviada) Publicado: Imprensa da Universidade de Oxford no Reino Unido. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. e Evans, David C., Currie, Phillip J., e Koppelhus, Eva (eds.). Parque Provincial dos Dinossauros: A Spectacular Ancient Ecosystem Revealed, Capítulo: Dinossauros Ornithischian. 2005, Publicado: Imprensa da Universidade de Indiana, em Bloomington. Páginas 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; e Dupuis, Kevin, 2007. Uma juvenil Parasaurolophus braincase do Parque Provincial dos Dinossauros, Alberta, com comentários sobre a ontogenia de crista no gênero. Journal of Vertebrate Paleontology, vol. 27, número 3 páginas 642-650.
- Currie, Phillip J; e Russell, Dale A. “Geographic and stratigraphic distribution of dinosaur remains” in Dinosaur Provincial Park, p. 553.
- Weishampel, David B. (1990). “Distribuição dos Dinossauros”, em The Dinosauria (1ª), pp. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; e Noto, Christopher R. (2004). “Dinosaur Distribution”, in The Dinosauria (2nd), pp. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, da Nova Acta Regia Societas Scientarum Upsaliensis, série 4, vol. 7, número 5. (alemão). Páginas 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, publicado pela Cassell Ltd. 1979, edição 5, em Londres. ISBN 0-304-52257-0 Página 883.
- Weishampel, David B. e Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) de Utah, do Journal of Paleontology, vol. 53, edição 6, páginas 1422-1427.
- Eberth, David A. 2005. “A geologia”, em Dinosaur Provincial Park, págs. 54-82.
- Braman, Dennis R., e Koppelhus, Eva B. 2005. “Palinomorfos Campanianos”, no Parque Provincial dos Dinossauros, págs. 101-130.
- Russell, Dale A. An Odyssey in Time: Os Dinossauros da América do Norte, 1989. Editora: NorthWord Press, em Minocqua, Wisconsin. ISBN 1-55971-038-1 Páginas 160-164.
- Bakker, Robert T. 1986. The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, publicado por William Morrow, em Nova Iorque. ISBN 0-8217-2859-8 Página 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: Uma Perspectiva Original e Convincente da Vida no Reino dos Dinossauros, capítulo: Hadrosaurídeas II. 1985. Publicado pela Crescent Books, em Nova York. Páginas 122-127. ISBN 0-517-468905
- 28,0 28,1 28,2 28,3 28,4 Evans, David C., 2006. Homologias da cavidade nasal e função da crista craniana em dinossauros lambeosaurinos, do Journal of Paleobiology, vol. 32, número 1, páginas 109-125.
- 29,0 29,1 29,2 29,3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs , do Journal of Paleobiology, volume 1, fascículo 1, páginas 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology, da Imprensa da Universidade de Chicago, na página 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, do Journal of Orion (Munique), vol. 2. pp. 525-532 (língua alemã).
- Sternberg, Charles M. 1935. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous, do Journal of the Canada Department of Mines Bulletin (Geological Series), volume 77, número 52, nas páginas 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Réptiles Ruling and their Relatives, publicado em 1945 pelo American Museum of Natural History, Man and Nature Publications, 14, em Nova York. Página 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas, do Journal of Jarbuch Naturwissenschaften, volume 12, número 36, nas páginas 709-716. (alemão) 1924.
- Osmólska, Halszka, 1979. Glândulas salinas nasais em dinossauros, do Journal of Acta Palaeontologica Polonica, vol. 24, páginas 205-215.
- 36.0 36.1 Ostrom, John H., 1962. As cristas cranianas dos dinossauros hadrosaurianos, do Journal of Postilla, vol. 62, pp. 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, publicado pela Master Books, em Green Forest. ISBN 0-89051-165-9 Página 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), do Journal of Paleobiology, vol. 7, número 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, do Journal of Paleontology, vol. 55, número 5, pp. 1046-1057.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur, dos Laboratórios Nacionais de Sandia (1997-12-05). Recuperado em 20 de Janeiro de 2009.
- Diegert, Carl F. e Williamson, Thomas E., 1998. Um modelo acústico digital do lambeosaurine hadrosaur Parasaurolophus tubicen do Journal of Vertebrate Paleontology, vol. 18, número 3, Suppl. p. 38A.
- Sullivan, Robert M. e Williamson, Thomas E., 1996. Um novo crânio de Parasaurolophus (forma cristalizada) do Novo México: características externas e internas (tomografias) e suas implicações funcionais, do Journal of Vertebrate Paleontology, vol. 16, número 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborar estrutura de resfriamento do SNC em grandes dinossauros Journal of Nature, vol. 275, nas pp. 441-443.
- Maryańska, Teresa e Osmólska, Halszka, em 1979. Aspectos da anatomia craniana hadrosauriana, do Journal of Lethaia, vol. 12, nas pp. 265-273.
- Cientistas usam paleontologia digital para produzir a voz do dinossauro Parasaurolophus; dos Laboratórios Nacionais Sandia.
- Restaurações de P. walkeri e P. cyrtocristatus, na mesma escala, de Scott Hartman; em Skeletal Drawing.com.
- Parasaurolophus página em Enchanted Learning.com