

Parasaurolophus (pronunțat /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, în mod obișnuit și /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-ə-LOH-fəs; însemnând „șopârlă cu creastă apropiată” cu referire la Saurolophus) este un gen de dinozaur ornithopod din perioada Cretacicului târziu din ceea ce este astăzi America de Nord, în urmă cu aproximativ 76-73 milioane de ani. Era un erbivor care mergea atât ca biped, cât și ca patruped. Sunt recunoscute trei specii: P. walkeri (specia tip), P. tubicen și P. cyrtocristatus cu creastă scurtă. Sunt cunoscute rămășițe din Alberta, Canada, și New Mexico și Utah, SUA. A fost descris pentru prima dată în 1922 de către William Parks, pornind de la un craniu și un schelet parțial din Alberta.
Parasaurolophus este un hadrosaurid, care face parte dintr-o familie diversă de dinozauri cretacici cunoscuți pentru gama lor de ornamente bizare ale capului. Acest gen este cunoscut pentru creasta sa craniană mare și elaborată, care, la cea mai mare dimensiune, formează un tub lung și curbat care se proiectează în sus și în spate din craniu. Charonosaurus din China, care este posibil să fi fost cea mai apropiată rudă a sa, avea un craniu asemănător și, potențial, o creastă similară. Creasta a fost mult discutată de oamenii de știință; consensul este că printre funcțiile majore se numără recunoașterea vizuală atât a speciei, cât și a sexului, rezonanța acustică și termoreglarea. Este unul dintre cei mai rari dinozauri de rață, fiind cunoscut doar dintr-o mână de exemplare bune.
Parasaurolophus se numără printre cei mai bine recunoscuți dinozauri și este frecvent portretizat în emisiuni de televiziune, filme, cărți, mărfuri și alte mijloace de comunicare legate de dinozauri. Alice Perry
Descriere

Mărimea lui Parasaurolophus walkeri în comparație cu cea a unui om.
Ca și în cazul majorității dinozaurilor, scheletul lui Parasaurolophus este incomplet cunoscut. Lungimea specimenului tip al lui P. walkeri este estimată la 9,5 metri (31 ft). Craniul său are o lungime de aproximativ 1,6 metri (5,2 ft), inclusiv creasta, în timp ce craniul tip al lui P. tubicen are o lungime de peste 2,0 metri (6,6 ft), ceea ce indică un animal mai mare. Greutatea sa este estimată la 2,5 tone (2,7 tone). Singurul său membru anterior cunoscut era relativ scurt pentru un hadrosaurid, cu o omoplat scurt, dar lat. Osul coapsei măsoară 103 centimetri (3,38 ft) la P. walkeri și este robust pentru lungimea sa, în comparație cu alți hadrosaurizi. Brațul superior și oasele pelviene erau, de asemenea, puternic construite.
Ca și alți hadrosaurizi, era capabil să meargă fie pe două picioare, fie pe patru. Probabil că a preferat să caute hrană pe patru picioare, dar a alergat pe două. Spinele neurale ale vertebrelor erau înalte, așa cum era obișnuit la lambeosaurine; mai înalte deasupra șoldurilor, ele creșteau înălțimea spatelui. Se cunosc amprente de piele pentru P. walkeri, care arată solzi uniformi, asemănători unor tuberculi, dar fără structuri mai mari.
Parasaurolophus walkeri cu detaliu de scalare.
Cea mai vizibilă trăsătură era creasta craniană, care ieșea din partea din spate a capului și era formată din premaxila și oasele nazale. Exemplarul tip P. walkeri are o crestătură în spinii neurali aproape de locul în care creasta s-ar lovi de spate, dar aceasta poate fi o patologie specifică acestui individ. William Parks, care a dat numele genului, a emis ipoteza că un ligament se întinde de la creastă la crestă până la crestătură pentru a susține capul. Deși ideea pare puțin probabilă, Parasaurolophus este uneori restaurat cu un lambou de piele de la creastă până la gât.
Creasta era goală, cu tuburi distincte care mergeau de la fiecare nară până la capătul crestei înainte de a inversa direcția și de a se îndrepta înapoi în josul crestei și în craniu. Tuburile erau cele mai simple la P. walkeri și mai complexe la P. tubicen, unde unele tuburi erau oarbe, iar altele se întâlneau și se separau. În timp ce P. walkeri și P. tubicen aveau creste lungi, cu o ușoară curbură, P. cyrtocristatus avea o creastă scurtă, cu un profil mai circular.
Clasificare
După cum sugerează și numele său, inițial s-a crezut că Parasaurolophus este înrudit îndeaproape cu Saurolophus din cauza crestei sale superficial asemănătoare. Cu toate acestea, a fost curând reevaluat ca membru al subfamiliei lambeosaurine a hadrosauridelor-Saurolophus este un hadrosaurin. De obicei, este interpretat ca o ramură separată a lambeosaurinelor, distinctă de Corythosaurus, Hypacrosaurus și Lambeosaurus, care au creastă de cască. Cea mai apropiată rudă cunoscută a sa pare a fi Charonosaurus, un lambeosaurin cu un craniu similar (dar fără creastă completă încă) din regiunea Amur din nord-estul Chinei, iar cei doi pot forma o cladă Parasaurolophini. P. cyrtocristatus, cu creasta sa scurtă și mai rotundă, poate fi cea mai bazală dintre cele trei specii cunoscute de Parasaurolophus, sau poate reprezenta exemplare subadulte sau femele ale lui P. tubicen.
Istorie
Descoperire și denumire
Castrul specimenului tip Parasaurolophus walkeri incomplet din Warszawa.
Semnând „șopârlă cu creastă apropiată”, numele Parasaurolophus derivă din grecescul para/παρα „lângă” sau „aproape”, saurus/σαυρος „șopârlă” și lophos/λοφος „creastă”. Se bazează pe ROM 768, un craniu și un schelet parțial căruia îi lipsește cea mai mare parte a cozii și picioarele posterioare de sub genunchi, care a fost găsit de o echipă de teren de la Universitatea din Toronto în 1920, în apropiere de Sand Creek, de-a lungul râului Red Deer din Alberta, Canada. Aceste roci sunt cunoscute în prezent sub numele de Formațiunea Dinosaur Park din Cretacicul superior de vârstă Campanian. William Parks a numit specimenul P. walkeri în onoarea lui Sir Byron Edmund Walker, președintele Consiliului de Administrație al Muzeului Regal din Ontario. Rămășițele de Parasaurolophus sunt rare în Alberta, existând doar un alt craniu parțial din (probabil) Formațiunea Dinosaur Park și trei specimene din Dinosaur Park fără craniu, care ar putea aparține genului. În unele liste de faună, există o mențiune cu privire la posibile materiale de P. walkeri în formațiunea Hell Creek din Montana, o unitate de rocă de vârstă Maastrichtian târziu. Această apariție nu este notată de Sullivan și Williamson în analiza lor din 1999 a genului și nu a fost dezvoltată în altă parte.
În 1921, Charles H. Sternberg a recuperat un craniu parțial (PMU.R1250) din ceea ce este acum cunoscut ca Formațiunea Kirtland, puțin mai tânără, din San Juan County, New Mexico. Acest specimen a fost trimis la Uppsala, Suedia, unde Carl Wiman l-a descris ca o a doua specie, P. tubicen, în 1931. Epitetul specific este derivat din latinescul tǔbǐcěn „trompetă”. Un al doilea craniu aproape complet de P. tubicen (NMMNH P-25100) a fost găsit în New Mexico în 1995. Folosind tomografia computerizată a acestui craniu, Robert Sullivan și Thomas Williamson au oferit genului un tratament monografic în 1999, acoperind aspecte ale anatomiei și taxonomiei sale, precum și funcțiile crestei sale. Williamson a publicat ulterior o analiză independentă a rămășițelor, nefiind de acord cu concluziile taxonomice.
John Ostrom a descris un alt specimen bun (FMNH P27393) din New Mexico ca fiind P. cyrtocristatus în 1961. Acesta include un craniu parțial cu o creastă scurtă, rotunjită, și o mare parte din scheletul postcranian, cu excepția picioarelor, a gâtului și a unor părți din coadă. Numele său specific derivă din latinescul curtus „scurtat” și cristatus „cu creastă”. Specimenul a fost găsit fie în partea superioară a Formațiunii Fruitland, fie, mai probabil, la baza Formațiunii Kirtland suprapuse. Aria de răspândire a acestei specii a fost extinsă în 1979, când David B. Weishampel și James A. Jensen au descris un craniu parțial cu o creastă similară (BYU 2467) din Formațiunea Kaiparowits din epoca Campanian din Garfield County, Utah. De atunci, un alt craniu a fost găsit în Utah cu morfologia crestei scurte/rotunde a lui P. cyrtocristatus.
Specie
Creier turnat al craniului lui Parasaurolophus walkeri, Muzeul de Istorie Naturală, Londra.
Gafă craniană Parasaurolophus cyrtocristatus, Field Museum of Natural History.
Specia tip P. walkeri, din Alberta, este cunoscută dintr-un singur exemplar. Se deosebește de P. tubicen prin faptul că are tuburi mai simple în creastă și de P. cyrtocristatus prin faptul că are o creastă lungă, nerotunjită și un braț superior mai lung decât antebrațul.
P. tubicen, din New Mexico, este cunoscută din rămășițele a cel puțin trei indivizi. Este cea mai mare specie, cu pasaje de aer mai complexe în creastă decât P. walkeri și cu o creastă mai lungă și mai dreaptă decât P. cyrtocristatus.
P. cyrtocristatus, din New Mexico și Utah, este cunoscut din trei exemplare posibile. Este cea mai mică specie, cu o creastă scurtă și rotunjită. Dimensiunile sale mici și forma crestei au determinat mai mulți cercetători să sugereze că reprezintă puii sau femelele lui P. tubicen, care provine aproximativ din aceeași perioadă și din aceeași formațiune din New Mexico. După cum a remarcat Thomas Williamson, materialul tip al lui P. cyrtocristatus are o dimensiune de aproximativ 72% din cea a lui P. tubicen, aproape de dimensiunea la care se interpretează că alte lambeosaurine încep să prezinte un dimorfism sexual definitiv în creasta lor (~70% din dimensiunea adultă). Această poziție a fost respinsă în recenziile recente ale lambeosaurinelor.
Paleobiologie
Paleoecologie
Gorgosaurus (stânga) atacând un Parasaurolophus cyrtocristatus.
Parasaurolophus walkeri, din Formațiunea Dinosaur Park, a făcut parte dintr-o faună diversă și bine documentată de animale preistorice, incluzând dinozauri bine cunoscuți precum Centrosaurus cu coarne, Styracosaurus și Chasmosaurus; colegii de rățoi Prosaurolophus, Gryposaurus, Corythosaurus și Lambeosaurus; tiranozaurul Gorgosaurus; precum și Edmontonia și Euoplocephalus cu armură. A fost un constituent rar al acestei faune. Formațiunea Dinosaur Park este interpretată ca un cadru cu relief scăzut de râuri și câmpii inundabile care au devenit mai mlăștinoase și au fost influențate de condițiile marine de-a lungul timpului, pe măsură ce Calea maritimă interioară vestică a transgresat spre vest. Clima era mai caldă decât în Alberta de astăzi, fără îngheț, dar cu anotimpuri mai umede și mai uscate. Coniferele erau, se pare, plantele dominante din coronament, cu un subarboret de ferigi, ferigi arboricole și angiosperme.
Speciile din Noul Mexic și-au împărțit mediul cu marele sauropod Alamosaurus, Kritosaurus cu cioc de rață, Pentaceratops cu coarne, Nodocephalosaurus cu armură, Saurornitholestes și tiranozauridele fără nume în prezent. Formațiunea Kirtland este interpretată ca fiind câmpii inundabile fluviale apărute după o retragere a căii maritime interioare vestice. Coniferele erau plantele dominante, iar dinozaurii cu coarne chasmosaurine erau aparent mai comuni decât hadrosauridele.
Alimentare
Ca hadrosaurid, Parasaurolophus era un ierbivor mare biped/quadruped, mâncând plante cu un craniu sofisticat care permitea o mișcare de măcinare analogă cu cea de mestecat. Dinții săi se înlocuiau continuu și erau împachetați în baterii dentare care conțineau sute de dinți, dintre care doar o mână relativă de dinți erau utilizați în orice moment. Își folosea ciocul pentru a recolta materialul vegetal, care era ținut în fălci de un organ asemănător obrazului. Hrănirea ar fi avut loc de la sol până la o înălțime de aproximativ 4 metri (13 ft). După cum a remarcat Bob Bakker, lambeosaurinele au ciocurile mai înguste decât hadrosaurinele, ceea ce implică faptul că Parasaurolophus și rudele sale puteau să se hrănească mai selectiv decât omologii lor cu cioc larg, fără creastă.
Creasta craniană
Au fost avansate multe ipoteze cu privire la funcțiile pe care le îndeplinea creasta craniană a lui Parasaurolophus, dar majoritatea au fost discreditate. În prezent se crede că este posibil să fi avut mai multe funcții: afișaj vizual pentru identificarea speciei și a sexului, amplificarea sunetului pentru comunicare și termoreglare. Nu este clar care a fost cea mai semnificativă și în ce momente din evoluția crestei și a pasajelor nazale interne ale acesteia.
Diferențe între specii și stadii de creștere
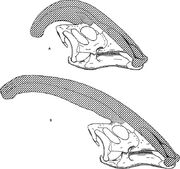
Diagrama care compară crestele nariale ale lui Parasaurolophus cyrtocristatus (a) și Parasaurolophus walkeri (b).
Ca și în cazul altor lambeosaurine, se crede că creasta craniană a lui Parasaurolophus s-a modificat odată cu vârsta și a fost o caracteristică dimorfică sexuală la adulți. James Hopson, unul dintre primii cercetători care a descris crestele lambeosaurinelor în termeni de astfel de distincții, a sugerat că P. cyrtocristatus, cu creasta sa mică, era forma feminină a lui P. tubicen. Thomas Williamson a sugerat că era forma juvenilă. Niciuna dintre cele două ipoteze nu a fost acceptată pe scară largă. Având în vedere că se cunosc doar șase cranii bune și o carcasă cerebrală juvenilă, materialul suplimentar va ajuta la clarificarea acestor relații potențiale. Williamson a remarcat că, în orice caz, Parasaurolophus juvenil avea probabil creste mici și rotunjite, precum P. cyrtocristatus, care probabil creșteau mai repede pe măsură ce indivizii se apropiau de maturitate. Restudierea recentă a unei creiere juvenile atribuite anterior lui Lambeosaurus, și acum atribuite lui Parasaurolophus, oferă dovezi că o creastă tubulară mică era prezentă la juvenili. Acest specimen păstrează o mică evazare în sus a oaselor frontale care era similară, dar mai mică decât cea observată la specimenele adulte; la adulți, frontalele formau o platformă care susținea baza crestei. Acest specimen indică, de asemenea, faptul că creșterea crestei la Parasaurolophus și profilul facial al indivizilor tineri diferă de modelul Corythosaurus-Hypacrosaurus-Lambeosaurus, în parte pentru că pe creasta lui Parasaurolophus lipsește „coxcomb-ul” osos subțire care alcătuiește partea superioară a crestei celorlalte trei lambeosaurine.
Ipoteze respinse privind funcția
Multe sugestii timpurii s-au concentrat pe adaptări pentru un stil de viață acvatic, urmând ipoteza că hadrosauridele erau amfibii, o linie de gândire comună până în anii 1960. Astfel, Alfred Sherwood Romer a propus că servea drept snorkel, Martin Wilfarth că era un accesoriu pentru o trompă mobilă folosită ca tub de respirație sau pentru a aduna hrană, Charles M. Sternberg că servea drept trapă de aer pentru a ține apa departe de plămâni, iar Ned Colbert că servea drept rezervor de aer pentru șederi prelungite sub apă.
Alte propuneri erau de natură mai fizică. După cum s-a menționat mai sus, William Parks a sugerat că era unită de vertebre cu ligamente sau mușchi și că ajuta la mișcarea și susținerea capului. Othenio Abel a propus că a fost folosit ca armă în luptă între membrii aceleiași specii, iar Andrew Milner a sugerat că ar putea fi folosit ca un deflector de frunziș, la fel ca și creasta de cască (numită „casque”) a casuarului. Alte propuneri au făcut din adăpostirea organelor specializate funcția principală. Halszka Osmólska a sugerat că adăpostea glandele saline, iar John Ostrom a sugerat că adăpostea zone extinse pentru țesutul olfactiv și un simț al mirosului mult îmbunătățit al lambeosaurinelor, care nu aveau capacități defensive evidente. O sugestie neobișnuită, făcută de creaționistul Duane Gish, este că creasta adăpostea glande chimice care îi permiteau să arunce jeturi de „foc” chimic asupra dușmanilor, similar cu gândacul bombardier din zilele noastre.
Majoritatea acestor ipoteze au fost discreditate sau respinse. De exemplu, nu există nicio gaură la capătul crestei pentru o funcție de scufundare. Nu există cicatrici musculare pentru o proboscisă și este îndoielnic faptul că un animal cu cioc ar avea nevoie de una. În calitate de sas propus, acesta nu ar fi ținut afară apa. Rezervorul de aer propus ar fi fost insuficient pentru un animal de mărimea lui Parasaurolophus. Alte hadrosauride aveau capete mari fără a avea nevoie de creste mari și goale care să servească drept puncte de fixare pentru ligamentele de susținere. De asemenea, niciuna dintre propuneri nu explică de ce creasta are o astfel de formă, de ce alte lambeosaurine ar trebui să aibă creste care arată mult diferit, dar îndeplinesc o funcție similară, cum se descurcau hadrosauridele fără creastă sau cu creastă solidă fără astfel de capacități sau de ce unele hadrosauride aveau creastă solidă. Aceste considerații au un impact deosebit asupra ipotezelor bazate pe creșterea capacităților sistemelor deja prezente în animal, cum ar fi ipoteza glandei saline și a olfacției, și indică faptul că acestea nu erau funcțiile primare ale crestei. În plus, lucrările asupra cavității nazale a lambeosaurinelor arată că nervii olfactivi și țesutul senzorial corespunzător se aflau în mare parte în afara porțiunii pasajelor nazale din creastă, astfel că extinderea crestei a avut puțin de-a face cu simțul mirosului.
Restaurarea vieții lui Parasaurolophus walkeri cu un ipotetic volan de piele și dungi pe creastă.
În schimb, funcțiile sociale și fiziologice au devenit mai susținute ca funcții ale crestei, concentrându-se pe identificarea și comunicarea vizuală și auditivă. Fiind un obiect mare, creasta are o valoare clară ca semnal vizual și diferențiază acest animal de contemporanii săi. Dimensiunea mare a orbitelor oculare ale hadrosauridelor și prezența inelelor sclerotice în ochi implică o vedere acută și obiceiuri diurne, dovadă că vederea era importantă pentru aceste animale. Dacă, așa cum este ilustrat în mod obișnuit, un volan de piele se întindea de la creastă până la gât sau spate, spectacolul vizual propus ar fi fost mult mai spectaculos. Așa cum sugerează și alte cranii de lambeosaurine, creasta lui Parasaurolophus a permis probabil atât identificarea speciei (cum ar fi separarea acestuia de Corythosaurus sau Lambeosaurus), cât și determinarea între masculi și femele, pe baza formei și a dimensiunii.
Funcția sonoră
Model computerizat al pasajelor nazale din creasta lui P. tubicen.
Cu toate acestea, aspectul exterior al crestei nu corespunde cu anatomia internă complexă a pasajelor nazale, ceea ce sugerează o altă funcție justificată de utilizarea spațiului intern. Carl Wiman a fost primul care a propus, în 1931, că pasajele îndeplineau o funcție de semnalizare auditivă, ca un crumhorn; Hopson și David B. Weishampel au revizuit această idee în anii 1970 și 1980. Hopson a constatat că există dovezi anatomice că hadrosauridele aveau un auz puternic. Există cel puțin un exemplu, la Corythosaurus, înrudit cu acesta, al unui staps subțire (osul urechii de reptilă) la locul lui, care, combinat cu un spațiu mare pentru timpan, implică o ureche medie sensibilă. În plus, lagena hadrosauridelor este alungită ca la crocodilieni, ceea ce indică faptul că porțiunea auditivă a urechii interne era bine dezvoltată. Weishampel a sugerat că P. walkeri era capabil să producă frecvențe cuprinse între 48 și 240 Hz, iar P. cyrtocristatus (interpretat ca o formă juvenilă de creastă) între 75 și 375 Hz. Bazându-se pe asemănarea urechilor interne ale hadrosauridelor cu cele ale crocodililor, el a propus, de asemenea, că hadrosauridele adulte erau sensibile la frecvențe înalte, cum ar fi cele pe care le-ar putea produce puii lor. Potrivit lui Weishampel, acest lucru este în concordanță cu comunicarea dintre părinți și urmași.
Modelarea computerizată a unui specimen bine conservat de P. tubicen, cu pasaje de aer mai complexe decât cele ale lui P. walkeri, a permis reconstrucția posibilului sunet produs de creasta sa. Calea principală rezonează la aproximativ 30 Hz, dar anatomia complicată a sinusurilor provoacă vârfuri și văi în sunet.
Funcția de răcire
Suprafața mare și vascularizarea crestei sugerează, de asemenea, o funcție de termoreglare. P.E. Wheeler a sugerat pentru prima dată această utilizare în 1978 ca o modalitate de a menține creierul rece. Teresa Maryańska și Osmólska au propus, de asemenea, termoreglarea cam în aceeași perioadă, iar Sullivan și Williamson s-au interesat în continuare. Discuția din 2006 a lui David Evans despre funcțiile crestei lambeosaurine a fost favorabilă ideii, cel puțin ca factor inițial pentru evoluția expansiunii crestei.
In The Media
Parasaurolophus este un dinozaur foarte popular. A apărut în filmul Disney’s Dinosaur ca membru al unei turme. De asemenea, a avut mai multe apariții în celebrul documentar Clash of the Dinosaurs. A apărut, de asemenea, în populara emisiune TV Prehistoric Park, unde a devenit prada carnivorelor mari Deinosuchus și Albertosaurus. A avut câteva apariții în filmele Jurassic Park, ca membru al turmei în primul film, fiind apoi ținut captiv de vânători în al doilea și fugind împreună cu Corythosaurus în al treilea. și personajul Dweeb în „We’re Back! A Dinosaur’s Story”, este el însuși un Parasaurolophus.
Parasaurolophus apare și în Turok, ca un mâncător de plante docil, care în mod normal nu este dăunător, dar poate fi agresiv dacă este provocat grav.
Există și un zord Parasaurolophus în Power Rangers: Dino Thunder și Power Rangers: Dino Charge.
Parasaurolophus apare pentru scurt timp la începutul filmului Disney Pixar The Good Dinosaur.
Vezi și
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, pagina 229. Publicat: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Chapter: Parasaurolophus, paginile 678-684. Publicat: 1997, McFarland & Co, în Jefferson, Carolina de Nord. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull și Wright, Hadrosaurian Dinosaurs of North America, pp. 209-213.
- Brett-Surman, Michael K. și Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, capitol: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, paginile 135-169. Publicat, 2006, Indiana University Press, în Bloomington și Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; și Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; și Dodson, Peter (eds.) The Dinosauria, ediția a 2-a, capitolul: Hadrosauridae, paginile 438-463. Publicat: 2004, University of California Press, în Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, volumul 13, paginile 1-32. Publicat: 1922, Studii ale Universității din Toronto, Seria Geologie.
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. și Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, din seria New Mexico Museum of Natural History and Science Bulletin, 15, paginile 1-52. Publicat: 1999, New Mexico Museum of Natural History and Science, în Albuqueque, New Mexico.
- 8.0 8.1 8.2 Ostrom, John H. 1961 O nouă specie de dinozaur hadrosaurian din Cretacicul din New Mexico, Journal of Paleontology, volumul 35, numărul 3, la paginile 575-577.
- Gilmore, Charles W., On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, volumul 38, numărul 43, paginile 29-48, Parks. Publicat: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. și Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; și Dodson, Peter (eds.) The Dinosauria, 1st edition, Chapter: Hadrosauridae, paginile 534-561. Publicat: 1990, University of California Press din Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., și Reisz, Robert R. 2007. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta, din Journal of Vertebrate Paleontology, vol. 27 numărul 2, la paginile 373-393.
- Godefroit, Pascal, Shuqin Zan; și Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaur din Maastrichtianul târziu din nord-estul Chinei, din Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, paginile 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Published by New Mexico Museum of Natural History and Science, in Albuqueque, New Mexico. Paginile 191-213.
- Liddell, Henry George și Robert Scott, 1980. A Greek-English Lexicon (A Greek-English Lexicon) (Ediție prescurtată) Publicat: Ed: Oxford University Press în Marea Britanie. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. și Evans, David C., Currie, Phillip J., și Koppelhus, Eva (eds.). Parcul Provincial Dinosaur: A Spectacular Ancient Ecosystem Revealed, Capitol: Ornithischian Dinosaurs. 2005, Publicat: Indiana University Press, în Bloomington. Paginile 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; și Dupuis, Kevin, 2007. O carcasă cerebrală juvenilă de Parasaurolophus din Dinosaur Provincial Park, Alberta, cu comentarii privind ontogenia crestei în gen. Journal of Vertebrate Paleontology, vol. 27, numărul 3, paginile 642-650.
- Currie, Phillip J; și Russell, Dale A. „Geographic and stratigraphic distribution of dinosaur remains” în Dinosaur Provincial Park, pag. 553.
- Weishampel, David B. (1990). „Dinosaur Distribution”, în The Dinosauria (1st), pp. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; și Noto, Christopher R. (2004). „Dinosaur Distribution”, în The Dinosauria (2nd), pp. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, din Nova Acta Regia Societas Scientarum Upsaliensis, seria 4, vol. 7, nr. 5. (în germană). Pagini 1-11.
- 21.0 21.1. Simpson, D.P. Cassell’s Latin Dictionary, publicat de Cassell Ltd. (în limba engleză). 1979, ediția 5, la Londra. ISBN 0-304-52257-0 Pagina 883.
- Weishampel, David B. și Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) din Utah, din Journal of Paleontology, vol. 53, nr. 6, pag. 1422-1427.
- Eberth, David A. 2005. „The geology”, în Dinosaur Provincial Park, în Dinosaur Provincial Park, pp. 54-82.
- Braman, Dennis R., și Koppelhus, Eva B. 2005. „Campanian palynomorphs”, în Dinosaur Provincial Park, pp. 101-130.
- Russell, Dale A. An Odyssey in Time: Dinosaurs of North America, 1989. Editor: NorthWord Press, în Minocqua, Wisconsin. ISBN 1-55971-038-1 Paginile 160-164.
- Bakker, Robert T. 1986. The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, publicat de William Morrow, în New York. ISBN 0-8217-2859-8 Pagina 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs (Enciclopedia ilustrată a dinozaurilor): An Original and Compelling Insight into Life in the Dinosaur Kingdom, cap.: „O perspectivă originală și convingătoare asupra vieții în regatul dinozaurilor”: Hadrosauridele II. 1985. Publicat de Crescent Books, în New York. Paginile 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs, din Journal of Paleobiology, vol. 32, issue 1, Pages 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs , din Journal of Paleobiology, volumul 1, numărul 1, paginile 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology, din University of Chicago Press, la pagina 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, din Journal of Orion (München), vol. 2. pp. 525-532 (în limba germană).
- Sternberg, Charles M. 1935. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous din Journal of the Canada Department of Mines Bulletin (Geological Series), volumul 77, numărul 52, la paginile 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, publicată în 1945 de Muzeul American de Istorie Naturală, Man and Nature Publications, 14, la New York. Pagina 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas, din revista Jarbuch Naturwissenschaften, volumul 12, numărul 36, la paginile 709-716. (germană) 1924.
- Osmólska, Halszka, 1979. Glandele saline nazale la dinozauri, din Revista Acta Palaeontologica Polonica, vol. 24, pag. 205-215.
- 36.0 36.1 Ostrom, John H., 1962. The cranial crests of hadrosaurian dinosaurs, din Journal of Postilla, vol. 62, pag. 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, publicat de Master Books, în Green Forest. ISBN 0-89051-165-9 Pagina 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), din Journal of Paleobiology, vol. 7, issue 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, din Journal of Paleontology, vol. 55, issue 5, pp. 1046-1057.
- Oamenii de știință folosesc paleontologia digitală pentru a produce vocea dinozaurului Parasaurolophus, de la Sandia National Laboratories (1997-12-05). Retrieved on January 20th, 2009.
- Diegert, Carl F. și Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen from the Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. și Williamson, Thomas E., 1996. A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications, din Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Structura elaborată de răcire a SNC la dinozaurii mari Journal of Nature, vol. 275, pe pp. 441-443.
- Maryańska, Teresa și Osmólska, Halszka, în 1979. Aspects of hadrosaurian cranial anatomy, din Journal of Lethaia, vol. 12, la pp. 265-273.
- Oamenii de știință folosesc paleontologia digitală pentru a produce vocea dinozaurului Parasaurolophus; de la Sandia National Laboratories.
- Restaurări ale lui P. walkeri și P. cyrtocristatus, la aceeași scară, de Scott Hartman; la Skeletal Drawing.com.
- Pagina despre Parasaurolophus de pe Enchanted Learning.com
.