

Parasaurolophus (uttalas /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, vanligen även /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; betyder ”ödla med nära kamm” med hänvisning till Saurolophus) är ett släkte av ornithopoddinosaurier från den sena kritaperioden i det som nu är Nordamerika, för cirka 76-73 miljoner år sedan. Det var en växtätare som gick både som tvåbent och fyrbent. Tre arter är kända: P. walkeri (typarten), P. tubicen och den kortkammade P. cyrtocristatus. Rester är kända från Alberta i Kanada samt New Mexico och Utah i USA. Den beskrevs första gången 1922 av William Parks från en skalle och ett partiellt skelett i Alberta.
Parasaurolophus är en hadrosaurid, en del av en varierad familj av dinosaurier från krittiden som är kända för sina olika bisarra huvudprydnader. Detta släkte är känt för sin stora, utarbetade kraniekista, som i sin största form bildar ett långt böjt rör som skjuter ut uppåt och bakåt från kraniet. Charonosaurus från Kina, som kan ha varit dess närmaste släkting, hade en liknande skalle och eventuellt en liknande kam. Forskarna har diskuterat kammen mycket, men de flesta är överens om att de viktigaste funktionerna var visuell igenkänning av både art och kön, akustisk resonans och värmereglering. Den är en av de mer sällsynta ankdjuren och är känd från endast en handfull bra exemplar.
Parasaurolophus är en av de mest välkända dinosaurierna och porträtteras ofta i dinosaurierelaterade tv-serier, filmer, böcker, legoartiklar och andra medier. Alice Perry
Beskrivning

Storleken på Parasaurolophus walkeri jämfört med en människa.
Som för de flesta dinosaurier är Parasaurolophus skelett ofullständigt känt. Längden på typexemplaret av P. walkeri uppskattas till 9,5 meter. Dess skalle är cirka 1,6 meter lång, inklusive kammen, medan typskallen av P. tubicen är över 2,0 meter lång, vilket tyder på ett större djur. Dess vikt uppskattas till 2,5 ton (2,7 ton). Dess enda kända framben var relativt kort för en hadrosaurid, med ett kort men brett skulderblad. Lårbenet mäter 103 centimeter hos P. walkeri och är robust för sin längd jämfört med andra hadrosaurider. Överarms- och bäckenbenen var också kraftigt byggda.
Likt andra hadrosaurider kunde den gå på antingen två eller fyra ben. Den föredrog förmodligen att söka föda på fyra ben, men sprang på två. Kotornas neuralryggar var höga, vilket var vanligt hos lambeosauriner; de var högst över höfterna och ökade ryggens höjd. Hudavtryck är kända för P. walkeri, som visar enhetliga tuberkelliknande fjäll men inga större strukturer.
Parasaurolophus walkeri med fjälldetalj.
Det mest påtagliga kännetecknet var kraniekammen, som stack ut från huvudets bakre del och som bestod av förkäke och näshuvudben. Typexemplaret av P. walkeri har en skåra i neuralryggarna nära den plats där kammen skulle träffa ryggen, men detta kan vara en patologi som är specifik för den här individen. William Parks, som namngav släktet, antog att ett ligament löpte från kammen till skåran för att stödja huvudet. Även om idén verkar osannolik, återställs Parasaurolophus ibland med en hudlapp från krönet till halsen.
Krönet var ihåligt, med distinkta rör som ledde från varje näsborre till krönets ände för att sedan vända riktning och gå tillbaka ner i krönet och in i skallen. Rören var enklast hos P. walkeri och mer komplexa hos P. tubicen, där vissa rör var blinda och andra möttes och skiljdes åt. Medan P. walkeri och P. tubicen hade långa kammar med endast svag krökning hade P. cyrtocristatus en kort kam med en mer cirkulär profil.
Klassificering
Som namnet antyder trodde man ursprungligen att Parasaurolophus var nära besläktad med Saurolophus på grund av dess ytligt sett liknande kam. Den omvärderades dock snart som en medlem av den lambeosauriska underfamiljen av hadrosaurider – Saurolophus är en hadrosaurin. Den brukar tolkas som en separat utlöpare av lambeosaurinerna, som skiljer sig från de hjälmkammade Corythosaurus, Hypacrosaurus och Lambeosaurus. Dess närmaste kända släkting tycks vara Charonosaurus, en lambeosaurin med ett liknande kranium (men ännu ingen komplett kam) från Amurregionen i nordöstra Kina, och de två kan bilda en klad Parasaurolophini. P. cyrtocristatus, med sin korta, rundare kam, kan vara den mest basala av de tre kända Parasaurolophus-arterna, eller så kan den representera subadulta eller kvinnliga exemplar av P. tubicen.
Historia
Upptäckt och namnsättning
Gjutning av det ofullständiga typexemplaret av Parasaurolophus walkeri i Warszawa.
Namnet Parasaurolophus, som betyder ”ödla med nära kamm”, härrör från grekiskans para/παρα ”bredvid” eller ”nära”, saurus/σαυρος ”ödla” och lophos/λοφος ”kamm”. Den är baserad på ROM 768, en skalle och ett partiellt skelett som saknar större delen av svansen och bakbenen under knäna, som hittades av en fältgrupp från Torontos universitet 1920 nära Sand Creek längs Red Deer River i Alberta, Kanada. Dessa stenar är nu kända som den övre krittiska Dinosaur Park-formationen från Campanian-åldern. William Parks namngav exemplaret P. walkeri för att hedra Sir Byron Edmund Walker, ordförande i styrelsen för Royal Ontario Museum. Parasaurolophus-rester är sällsynta i Alberta, med endast ett annat partiellt kranium från (troligen) Dinosaur Park-formationen, och tre Dinosaur Park-exemplar utan kranium, som möjligen tillhör släktet. I vissa faunalistor nämns ett möjligt P. walkeri-material i Hell Creek-formationen i Montana, en bergart av sen maastrikisk ålder. Denna förekomst noteras inte av Sullivan och Williamson i deras genomgång av släktet från 1999, och har inte heller utvecklats ytterligare någon annanstans.
År 1921 återfann Charles H. Sternberg ett partiellt kranium (PMU.R1250) från vad som nu är känt som den något yngre Kirtlandformationen i San Juan County, New Mexico. Exemplaret skickades till Uppsala, Sverige, där Carl Wiman 1931 beskrev det som en andra art, P. tubicen. Det specifika epitetet kommer från latinets tǔbǐcěn ”trumpetare”. Ett andra, nästan komplett kranium av P. tubicen (NMMNH P-25100) hittades i New Mexico 1995. Med hjälp av datortomografi av detta kranium gav Robert Sullivan och Thomas Williamson släktet en monografisk behandling 1999, som omfattade aspekter av dess anatomi och taxonomi samt kammarens funktioner. Williamson publicerade senare en oberoende granskning av kvarlevorna, där han inte höll med om de taxonomiska slutsatserna.
John Ostrom beskrev 1961 ett annat bra exemplar (FMNH P27393) från New Mexico som P. cyrtocristatus. Det omfattar en del av skallen med en kort, rundad kam och stora delar av det postkraniella skelettet utom fötterna, nacken och delar av svansen. Dess specifika namn kommer från latinets curtus ”förkortad” och cristatus ”krönt”. Exemplaret hittades antingen i toppen av Fruitlandformationen eller, mer troligt, i basen av den överliggande Kirtlandformationen. Artens utbredningsområde utvidgades 1979, då David B. Weishampel och James A. Jensen beskrev ett partiellt kranium med en liknande kam (BYU 2467) från Kaiparowits-formationen från den kampaniska åldern i Garfield County, Utah. Sedan dess har ytterligare ett kranium hittats i Utah med den korta/runda P. cyrtocristatus crest morfologi.
Species
Parasaurolophus walkeri kraniumgjutning, Natural History Museum, London.
Parasaurolophus cyrtocristatus kranieavgjutning, Field Museum of Natural History.
Typarten P. walkeri, från Alberta, är känd från ett enda exemplar. Den skiljer sig från P. tubicen genom att ha enklare rör i kammen, och från P. cyrtocristatus genom att ha en lång, oavrundad kam och en längre överarm än underarm.
P. tubicen, från New Mexico, är känd från kvarlevorna av minst tre individer. Det är den största arten, med mer komplexa luftpassager i krönet än P. walkeri, och ett längre och rakare krön än P. cyrtocristatus.
P. cyrtocristatus, från New Mexico och Utah, är känd från tre möjliga exemplar. Det är den minsta arten, med en kort rundad kam. Dess lilla storlek och kammarens form har fått flera forskare att föreslå att den representerar juveniler eller honor av P. tubicen, som är från ungefär samma tid och från samma formation i New Mexico. Som Thomas Williamson noterade är typmaterialet av P. cyrtocristatus ungefär 72 % av P. tubicens storlek, nära den storlek vid vilken andra lambeosauriner tolkas börja visa definitiv könsdimorfism i sina kammar (~70 % av vuxenstorlek). Denna ståndpunkt har förkastats i nyligen gjorda granskningar av lambeosauriner.
Paleobiology
Paleoecology
Gorgosaurus (till vänster) attackerar en Parasaurolophus cyrtocristatus.
Parasaurolophus walkeri, från Dinosaur Park-formationen, ingick i en mångsidig och väldokumenterad fauna av förhistoriska djur, inklusive välkända dinosaurier som den horniga Centrosaurus, Styracosaurus och Chasmosaurus; andfåglar som Prosaurolophus, Gryposaurus, Corythosaurus och Lambeosaurus, tyrannosaurid Gorgosaurus och pansrade Edmontonia och Euoplocephalus. Den var en sällsynt beståndsdel i denna fauna. Dinosaur Park-formationen tolkas som en lågreliefmiljö av floder och flodslätter som blev mer sumpiga och påverkade av marina förhållanden med tiden när den västra inre sjövägen gick västerut. Klimatet var varmare än i dagens Alberta, utan frost, men med både våtare och torrare årstider. Barrträd var tydligen de dominerande växterna i trädkronorna, med en undervegetation av ormbunkar, trädfarna och angiospermer.
De nymexikanska arterna delade sin miljö med de stora sauropoderna Alamosaurus, Kritosaurus med ankben, den horniga Pentaceratops, den pansarförsedda Nodocephalosaurus, Saurornitholestes och för närvarande icke namngivna tyrannosaurider. Kirtlandformationen tolkas som flodflodslätter som uppstod efter att den västra inre sjövägen dragit sig tillbaka. Barrträd var de dominerande växterna, och chasmosauriska horndinosaurier var tydligen vanligare än hadrosaurider.
Föda
Som hadrosaurid var Parasaurolophus en stor tvåbent/kvadbent växtätare, som åt växter med en sofistikerad skalle som tillät en slipande rörelse som liknade tuggning. Dess tänder byttes ständigt ut och packades i tandbatterier som innehöll hundratals tänder, varav endast en relativ handfull var i bruk vid varje tidpunkt. Den använde sin näbb för att skörda växtmaterial, som hölls i käkarna av ett kindliknande organ. Födosökandet kan ha skett från marken upp till cirka 4 meters höjd. Som Bob Bakker påpekat har lambeosauriner smalare näbbar än hadrosauriner, vilket innebär att Parasaurolophus och dess släktingar kunde livnära sig mer selektivt än sina brednäbbade, kamlösa motsvarigheter.
Kraniekista
Många hypoteser har framförts om vilka funktioner kraniekistan hos Parasaurolophus fyllde, men de flesta har misskrediterats. Man tror nu att den kan ha haft flera funktioner: visuell visning för att identifiera art och kön, ljudförstärkning för kommunikation och termoreglering. Det är oklart vilken som hade störst betydelse vid vilka tidpunkter i utvecklingen av kammen och dess inre näsgångar.
Differenser mellan arter och tillväxtstadier
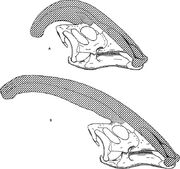
Diagram som jämför nariakammarna hos Parasaurolophus cyrtocristatus (a) och Parasaurolophus walkeri (b).
Som för andra lambeosauriner tror man att kraniekistan hos Parasaurolophus förändrades med åldern och var ett könsdimorft kännetecken hos vuxna. James Hopson, en av de första forskarna som beskrev lambeosaurinernas kammar i termer av sådana skillnader, föreslog att P. cyrtocristatus, med sin lilla kam, var den kvinnliga formen av P. tubicen. Thomas Williamson föreslog att det var den unga formen. Ingen av hypoteserna blev allmänt accepterad. Eftersom endast sex goda kranier och en juvenil hjärnkropp är kända, kommer ytterligare material att bidra till att klargöra dessa potentiella släktskap. Williamson noterade att i vilket fall som helst hade juvenila Parasaurolophus troligen små, rundade kammar som P. cyrtocristatus, som troligen växte snabbare när individerna närmade sig sin mognad. Nyligen genomfördes en ny undersökning av en juvenil hjärnkropp som tidigare tilldelats Lambeosaurus, och som nu tilldelats Parasaurolophus, vilket ger bevis för att en liten rörformig kam fanns hos juvenila djur. Detta exemplar bevarar en liten uppåtriktad utbuktning av frontalbenen som liknade men var mindre än vad som ses hos vuxna exemplar; hos vuxna bildade frontalbenen en plattform som stödde kammarens bas. Detta exemplar visar också att kammarens tillväxt hos Parasaurolophus och ansiktsprofilen hos unga individer skiljde sig från Corythosaurus-Hypacrosaurus-Lambeosaurus-modellen, delvis på grund av att kammen hos Parasaurolophus saknar den tunna beniga ”kaxkam” som utgör den övre delen av kammen hos de andra tre lambeosaurinerna.
Förkastade hypoteser om funktion
Många tidiga förslag fokuserade på anpassningar för en vattenlevande livsstil, i enlighet med hypotesen att hadrosaurider var amfibiska, en vanlig tankelinje fram till 1960-talet. Sålunda föreslog Alfred Sherwood Romer att den fungerade som en snorkel, Martin Wilfarth att den var ett fäste för en rörlig proboscis som användes som andningsrör eller för att samla mat, Charles M. Sternberg att den fungerade som en luftfälla för att hålla vatten borta från lungorna, och Ned Colbert att den fungerade som en luftreservoar för långvariga vistelser under vattnet.
Andra förslag var mer fysiska till sin natur. Som nämnts ovan föreslog William Parks att den var sammanfogad med kotorna med hjälp av ligament eller muskler och att den hjälpte till att flytta och stödja huvudet. Othenio Abel föreslog att den användes som ett vapen i strid mellan medlemmar av samma art, och Andrew Milner föreslog att den kunde användas som en lövavledare, likt hjälmkammen (kallad ”casque”) hos kasuaren. I ytterligare andra förslag var den viktigaste funktionen att hysa specialiserade organ. Halszka Osmólska föreslog att den innehöll saltkörtlar, och John Ostrom föreslog att den innehöll utökade områden för luktvävnad och ett mycket förbättrat luktsinne hos lambeosaurinerna, som inte hade någon uppenbar försvarsförmåga. Ett ovanligt förslag, som kreationisten Duane Gish har lagt fram, är att kammen hyste kemiska körtlar som gjorde det möjligt för den att kasta strålar av kemisk ”eld” mot fiender, i likhet med dagens bombardierbagge.
De flesta av dessa hypoteser har misskrediterats eller förkastats. Det finns till exempel inget hål i slutet av krönet för en snorkelfunktion. Det finns inga muskelärr för en proboscis och det är tveksamt att ett djur med näbb skulle behöva en sådan. Som en föreslagen luftsluss skulle den inte ha hållit vatten ute. Den föreslagna luftbehållaren skulle ha varit otillräcklig för ett djur av Parasaurolophus storlek. Andra hadrosaurider hade stora huvuden utan att behöva stora ihåliga kammar som skulle fungera som fästpunkter för stödjande ligament. Inget av förslagen förklarar heller varför krönet har en sådan form, varför andra lambeosauriner skulle ha krön som ser mycket annorlunda ut men fyller en liknande funktion, hur kamlösa eller hadrosaurider med fasta krön klarade sig utan sådana funktioner, eller varför vissa hadrosaurider hade fasta krön. Dessa överväganden påverkar särskilt hypoteser som bygger på att öka kapaciteten hos system som redan finns i djuret, t.ex. saltkörteln och lukthypoteserna, och visar att dessa inte var kammarens primära funktioner. Dessutom visar arbete med näshålan hos lambeosauriner att luktnerver och motsvarande sensorisk vävnad till stor del låg utanför den del av näsgångarna som fanns i kammen, så kammens utvidgning hade inte mycket att göra med luktsinnet.
Sociala funktioner
Livsrestaurering av Parasaurolophus walkeri med hypotetisk hudfrilla och ränder på krönet.
Istället har sociala och fysiologiska funktioner fått mer stöd som funktion(er) för krönet, med fokus på visuell och auditiv identifiering och kommunikation. Som ett stort objekt har kammen ett tydligt värde som en visuell signal och skiljer detta djur från sina jämnåriga artfränder. Den stora storleken på hadrosauridernas ögonhålor och förekomsten av sklerotiska ringar i ögonen antyder akut syn och dygnsvanor, vilket bevisar att synen var viktig för dessa djur. Om, som ofta illustreras, en hudkrans sträckte sig från kammen till nacken eller ryggen, skulle den föreslagna visuella visningen ha varit mycket mer prålig. Som föreslås av andra lambeosauriska kranier, möjliggjorde kammen hos Parasaurolophus troligen både artbestämning (som att skilja den från Corythosaurus eller Lambeosaurus) och bestämning mellan hanar och honor, baserat på form och storlek.
Ljudfunktion
Datormodell av näsgångarna i kammen hos P. tubicen.
Det yttre utseendet på kammen motsvarar dock inte den komplexa inre anatomin hos näsgångarna, vilket tyder på att en annan funktion stod för användningen av det inre utrymmet. Carl Wiman var den förste som 1931 föreslog att passagerna hade en auditiv signalfunktion, likt ett krummhorn. Hopson och David B. Weishampel återkom till denna idé på 1970- och 1980-talen. Hopson fann att det finns anatomiska bevis för att hadrosaurider hade en stark hörsel. Det finns åtminstone ett exempel, hos den besläktade Corythosaurus, på ett smalt stapes (reptilöronben) på plats, vilket i kombination med ett stort utrymme för trumhinnan antyder ett känsligt mellanöra. Dessutom är hadrosauridens lagena långsträckt som hos en krokodil, vilket tyder på att den auditiva delen av innerörat var välutvecklad. Weishampel föreslog att P. walkeri kunde producera frekvenser på 48 till 240 Hz och P. cyrtocristatus (tolkad som en juvenil kamform) 75 till 375 Hz. Baserat på likheten mellan hadrosauridernas inneröron och krokodilernas föreslog han också att vuxna hadrosaurider var känsliga för höga frekvenser, sådana som deras avkomma skulle kunna producera. Enligt Weishampel är detta förenligt med att föräldrar och avkomma kommunicerar.
Datormodellering av ett välbevarat exemplar av P. tubicen, med mer komplexa luftkanaler än hos P. walkeri, har gjort det möjligt att rekonstruera det möjliga ljud som dess kam producerade. Huvudvägen resonerar vid cirka 30 Hz, men den komplicerade sinusanatomin orsakar toppar och dalar i ljudet.
Kylfunktion
Den stora ytan och vaskulariseringen av krönet tyder också på en termoreglerande funktion. P.E. Wheeler föreslog först denna användning 1978 som ett sätt att hålla hjärnan kall. Teresa Maryańska och Osmólska föreslog också termoreglering ungefär samtidigt, och Sullivan och Williamson intresserade sig vidare. David Evans diskussion 2006 om lambeosauriernas kamfunktion var gynnsam för idén, åtminstone som en inledande faktor för utvecklingen av kamutvidgning.
I media
Parasaurolophus är en mycket populär dinosaurie. Den var med i filmen Disneys Dinosaurier som en flockmedlem. Den gjorde också flera framträdanden i den berömda dokumentären Clash of the Dinosaurs. Den dök också upp i den populära TV-serien Prehistoric Park, där den blev byte för de stora köttätarna Deinosuchus och Albertosaurus. Den gjorde ett par framträdanden i Jurassic Park-filmerna, som en flockmedlem i den första filmen, sedan hölls den fången av jägare i den andra och sprang tillsammans med Corythosaurus i den tredje. och karaktären Dweeb i ”We’re Back! A Dinosaur’s Story” är själv en Parasaurolophus.
Parasaurolophus förekommer också i Turok, som en foglig växtätare som normalt sett inte är skadlig, men som kan vara aggressiv om den provoceras allvarligt.
Det finns också en Parasaurolophus zord i Power Rangers: Dino Thunder och Power Rangers: Dino Charge.
Parasaurolophus dyker upp kort i början av Disney Pixar-filmen The Good Dinosaur.
Se även
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurier Dinosaurier i Nordamerika, sid 229. Publicerad: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, kapitel: Parasaurolophus, sidorna 678-684. Publicerad: 1997, McFarland & Co, i Jefferson, North Carolina. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull och Wright, Hadrosaurian Dinosaurs of North America, s. 209-213.
- Brett-Surman, Michael K. och Wagner, Jonathan R. Carpenter, Kenneth (red.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, chapter: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, pages 135-169. Publicerad 2006, Indiana University Press, i Bloomington och Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; och Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; och Dodson, Peter (eds.) The Dinosauria, 2nd edition, chapter: Hadrosauridae, sidorna 438-463. Publicerad: 2004, University of California Press, i Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, volume 13, pages 1-32. Publicerad: 1922, University of Toronto Studies, Geology Series.
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. och Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, från serien New Mexico Museum of Natural History and Science Bulletin, 15, pages 1-52. Publicerad: 1999, New Mexico Museum of Natural History and Science, i Albuqueque, New Mexico.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, Journal of Paleontology, Volume 35, 3rd issue, on pages 575-577.
- Gilmore, Charles W., On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, volume 38, issue 43, pages 29-48, Parks. Publicerad: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. and Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) The Dinosauria, 1st edition, Chapter: Hadrosauridae, sidorna 534-561. Publicerad: 1990, University of California Press i Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C. och Reisz, Robert R. 2007. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta, från Journal of Vertebrate Paleontology, vol. 27 issue 2, på sidorna 373-393.
- Godefroit, Pascal, Shuqin Zan och Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaur från det sena Maastrichtian i nordöstra Kina, från Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, pages 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Published by New Mexico Museum of Natural History and Science, in Albuqueque, New Mexico. Sidorna 191-213.
- Liddell, Henry George och Robert Scott, 1980. A Greek-English Lexicon (förkortad utgåva) Publicerad: Oxford University Press i Storbritannien. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. and Evans, David C., Currie, Phillip J., and Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, kapitel: Ornithischian Dinosaurs. 2005, Publicerad: Indiana University Press, i Bloomington. Sidorna 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R. och Dupuis, Kevin, 2007. En juvenil hjärnkropp av Parasaurolophus från Dinosaur Provincial Park, Alberta, med kommentarer om kammarens ontogeni i släktet. Journal of Vertebrate Paleontology, vol. 27, issue 3 pages 642-650.
- Currie, Phillip J; och Russell, Dale A. ”Geographic and stratigraphic distribution of dinosaur remains” i Dinosaur Provincial Park, s. 553.
- Weishampel, David B. (1990). ”Dinosauriernas utbredning”, i The Dinosauria (1st), s. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). ”Dinosauriernas utbredning”, i The Dinosauria (2nd), s. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, från Nova Acta Regia Societas Scientarum Upsaliensis, series 4, vol. 7, issue 5. (tyska). Sidorna 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, utgiven av Cassell Ltd. 1979, utgåva 5, i London. ISBN 0-304-52257-0 Sidan 883.
- Weishampel, David B. och Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) from Utah, från Journal of Paleontology, vol. 53, issue 6, pages 1422-1427.
- Eberth, David A. 2005. ”The geology”, i Dinosaur Provincial Park, s. 54-82.
- Braman, Dennis R., och Koppelhus, Eva B. 2005. ”Campanian palynomorphs”, i Dinosaur Provincial Park, s. 101-130.
- Russell, Dale A. An Odyssey in Time: Dinosaurs of North America, 1989. Förlag: NorthWord Press, i Minocqua, Wisconsin. ISBN 1-55971-038-1 Sidorna 160-164.
- Bakker, Robert T. 1986. The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, utgiven av William Morrow i New York. ISBN 0-8217-2859-8 Sidan 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom, kapitel: Hadrosaurider II. 1985. Publicerad av Crescent Books i New York. Sidorna 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs, från Journal of Paleobiology, vol. 32, issue 1, Pages 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs , från Journal of Paleobiology, volym ett, nummer ett, sidorna 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology , från University of Chicago Press, på sidan 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, från Journal of Orion (München), vol. 2. s. 525-532 (på tyska).
- Sternberg, Charles M. 1935. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous från Journal of the Canada Department of Mines Bulletin (Geological Series), volym 77, nummer 52, på sidorna 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, utgiven 1945 av American Museum of Natural History, Man and Nature Publications, 14, i New York. Sidan 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas från Journal of Jarbuch Naturwissenschaften, volym 12, nummer 36, på sidorna 709-716. (Tyska) 1924.
- Osmólska, Halszka, 1979. Nasala saltkörtlar hos dinosaurier, från tidskriften Acta Palaeontologica Polonica, vol. 24, sidorna 205-215.
- 36.0 36.1 Ostrom, John H., 1962. The cranial crests of hadrosaurian dinosaurs, från Journal of Postilla, vol. 62, pages 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, utgiven av Master Books, i Green Forest. ISBN 0-89051-165-9 Sida 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), från Journal of Paleobiology, vol. 7, issue 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, från Journal of Paleontology, vol. 55, issue 5, pp. 1046-1057.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur, från Sandia National Laboratories (1997-12-05). Hämtad den 20 januari 2009.
- Diegert, Carl F. och Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen from the Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. s. 38A.
- Sullivan, Robert M. och Williamson, Thomas E., 1996. A new skull of Parasaurolophus (long-cested form) from New Mexico: external and internal (CT scans) features and their functional implications, från Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp. 68A.
- Wheeler, P.E., 1978. Elaborate CNS cooling structure in large dinosaurs Journal of Nature, vol. 275, on pp. 441-443.
- Maryańska, Teresa och Osmólska, Halszka, 1979. Aspects of hadrosaurian cranial anatomy, från Journal of Lethaia, vol. 12, på sidorna 265-273.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur; från Sandia National Laboratories.
- Restaureringar av P. walkeri och P. cyrtocristatus, i samma skala, av Scott Hartman; på Skeletal Drawing.com.
- Parasaurolophus-sidan på Enchanted Learning.com
.