

Parasaurolophus (lausutaan /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, yleisesti myös /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; tarkoittaen Saurolophusiin viitaten ”lähellä harjaa oleva lisko”) on ornithopodi-dinosaurusten suku, joka on kotoisin myöhäisliitukaudelta nykyisen Pohjois-Amerikan alueelta noin 76-73 miljoonaa vuotta sitten. Se oli kasvinsyöjä, joka käveli sekä kaksi- että nelijalkaisena. Tunnettuja lajeja on kolme: P. walkeri (tyyppilaji), P. tubicen ja lyhytkarvainen P. cyrtocristatus. Muinaisjäännöksiä tunnetaan Albertasta, Kanadasta, sekä New Mexicosta ja Utahista, Yhdysvalloista. William Parks kuvasi sen ensimmäisen kerran vuonna 1922 Albertassa sijaitsevasta kallosta ja osittaisesta luurangosta.
Parasaurolophus on hadrosauridi, joka kuuluu moninaiseen liitukauden dinosaurusten perheeseen, joka tunnetaan erilaisista omituisista pääkoristeistaan. Tämä suku tunnetaan suuresta, taidokkaasta kallonharjasta, joka suurimmillaan muodostaa pitkän kaarevan putken, joka työntyy kallosta ylöspäin ja taaksepäin. Kiinasta peräisin olevalla Charonosauruksella, joka saattoi olla sen lähin sukulainen, oli samankaltainen kallo ja mahdollisesti samanlainen otsaharja. Tutkijat ovat keskustelleet paljon harjanteesta; yksimielisyys vallitsee siitä, että sen tärkeimpiä tehtäviä olivat lajin ja sukupuolen visuaalinen tunnistaminen, akustinen resonanssi ja lämmönsäätely. Se on yksi harvinaisemmista sorsalinnuista, sillä se tunnetaan vain muutamasta hyvästä yksilöstä.
Parasaurolophus on yksi tunnetuimmista dinosauruksista, ja sitä kuvataan usein dinosauruksiin liittyvissä televisio-ohjelmissa, elokuvissa, kirjoissa, tuotteissa ja muussa mediassa. Alice Perry
Kuvaus

Parasaurolophus walkerin koko verrattuna ihmiseen.
Kuten useimpien dinosaurusten kohdalla, Parasaurolophuksen luuranko tunnetaan puutteellisesti. P. walkeri -tyypin yksilön pituudeksi on arvioitu 9,5 metriä. Sen kallo on noin 1,6 metriä pitkä harja mukaan luettuna, kun taas P. tubicenin tyyppikallo on yli 2,0 metriä pitkä, mikä viittaa suurempaan eläimeen. Sen painoksi on arvioitu 2,5 tonnia. Sen ainoa tunnettu eturaaja oli hadrosauridiksi suhteellisen lyhyt, ja sen olkavarsi oli lyhyt mutta leveä. P. walkerin reisiluu on 103 senttimetriä pitkä ja muihin hadrosaurideihin verrattuna vankka pituuteensa nähden. Myös olkavarren ja lantion luut olivat järeitä.
Kuten muutkin hadrosauridit, se pystyi kävelemään joko kahdella tai neljällä jalalla. Se etsi todennäköisesti mieluummin ruokaa neljällä jalalla, mutta juoksi kahdella jalalla. Selkärangan neuraaliset selkärangat olivat korkeat, kuten lambeosaurineilla oli tapana; korkeimmillaan lantion yläpuolella ne lisäsivät selän korkeutta. P. walkerista tunnetaan ihopainanteita, joissa näkyy yhtenäisiä mukulamaisia suomuja, mutta ei suurempia rakenteita.
Parasaurolophus walkeri suomutuksen yksityiskohdilla.
Huomattavin piirre oli takaraivosta esiin työntyvä kallonharja, joka koostui premaxillasta ja nenäluista. P. walkeri -tyypin yksilössä on hermosäikeissä lovi lähellä kohtaa, jossa kallonharja osuisi selkään, mutta tämä saattaa olla tälle yksilölle ominainen patologia. William Parks, joka nimesi suvun, esitti hypoteesin, että nivelside kulkee harjanteelta lovelle ja tukee päätä. Vaikka ajatus vaikuttaa epätodennäköiseltä, Parasaurolophus on joskus restauroitu siten, että harjanteelta kaulalle ulottuu iholäppä.
Harjanne oli ontto, ja kummastakin sieraimesta johti erilliset putket harjanteen päähän, ennen kuin ne käänsivät suunnan ja suuntautuivat takaisin harjannetta alaspäin ja kalloon. Putket olivat yksinkertaisimmat P. walkerilla ja monimutkaisemmat P. tubicenilla, jossa jotkut putket olivat sokeita ja toiset kohtasivat ja erottuivat toisistaan. Kun P. walkerilla ja P. tubicenilla oli pitkät harjanteet, joissa oli vain vähäinen kaarevuus, P. cyrtocristatuksella oli lyhyt harjanne, jossa oli pyöreämpi profiili.
Luokittelu
Nimensä mukaisesti Parasaurolophus luultiin alun perin läheiseksi sukulaisuudeksi Saurolophuksen kanssa sen pinnallisesti samankaltaisen harjanteen vuoksi. Se arvioitiin kuitenkin pian uudelleen hadrosauriinien lambeosauriinien alaheimon jäseneksi – Saurolophus on hadrosauriini. Se tulkitaan yleensä lambeosauriinien erilliseksi sivuhaaraksi, joka eroaa kypäräpäisistä Corythosauruksesta, Hypacrosauruksesta ja Lambeosauruksesta. Sen lähin tunnettu sukulainen näyttää olevan Charonosaurus, lambeosauriini, jolla on samanlainen kallo (mutta ei vielä täydellistä kypärää) Koillis-Kiinan Amurin alueelta, ja nämä kaksi saattavat muodostaa Parasaurolophini-klaanin. P. cyrtocristatus, jolla on lyhyt, pyöreämpi harja, voi olla basaalisin kolmesta tunnetusta Parasaurolophus-lajista, tai se voi edustaa P. tubicenin subadultti- tai naarasyksilöitä.
Historia
Löytäminen ja nimeäminen
Puutteellisen Parasaurolophus walkeri -tyypin yksilön valukappale Warszawassa.
Nimi Parasaurolophus tarkoittaa ”lähellä olevaa harjaliskoa”, ja se on johdettu kreikan kielen sanoista para/παρα ”vieressä” tai ”lähellä”, saurus/σαυρος ”lisko” ja lophos/λοφος ”harja”. Se perustuu ROM 768 -nimiseen kalloon ja osittaiseen luurankoon, josta puuttuu suurin osa hännästä ja takajalat polvien alapuolelta ja jonka Toronton yliopiston kenttäryhmä löysi vuonna 1920 Sand Creekin läheltä Red Deer -joen varrella Albertassa Kanadassa. Nämä kivilajit tunnetaan nykyään kampanjakautisena ylemmän liitukauden Dinosaur Park -muodostumana. William Parks nimesi yksilön P. walkeri kuninkaallisen Ontarion museon johtokunnan puheenjohtajan Sir Byron Edmund Walkerin kunniaksi. Parasaurolophuksen jäännökset ovat Albertassa harvinaisia, ja vain yksi toinen osittainen kallo on (todennäköisesti) Dinosaur Park -muodostumasta, ja kolmesta Dinosaur Park -näytteestä puuttuu kallo, joka mahdollisesti kuuluu sukuun. Joissakin faunaluetteloissa on maininta mahdollisesta P. walkeri -aineistosta Montanan Hell Creek -muodostumassa, joka on myöhäismaastrichti-ikäinen kivilaji. Sullivan ja Williamson eivät mainitse tätä esiintymää vuoden 1999 katsauksessaan suvusta, eikä sitä ole käsitelty tarkemmin muualla.
Vuonna 1921 Charles H. Sternberg löysi osittaisen kallon (PMU.R1250) New Mexicon San Juanin piirikunnasta, New Mexicossa sijaitsevasta, nykyisin hieman nuoremmasta Kirtlandin muodostumasta. Tämä yksilö lähetettiin Uppsalaan, Ruotsiin, jossa Carl Wiman kuvasi sen toisena lajina, P. tubicen, vuonna 1931. Spesifinen epiteetti on johdettu latinankielisestä tǔbǐcěn ”trumpetisti”. Toinen, lähes täydellinen P. tubicen -kallo (NMMNH P-25100) löydettiin New Mexicosta vuonna 1995. Tämän kallon tietokonetomografian avulla Robert Sullivan ja Thomas Williamson antoivat suvulle monografisen käsittelyn vuonna 1999, jossa käsiteltiin sen anatomian ja taksonomian näkökohtia sekä harjan toimintoja. Williamson julkaisi myöhemmin jäännöksistä riippumattoman katsauksen, jossa hän oli eri mieltä taksonomisten johtopäätösten kanssa.
John Ostrom kuvasi toisen hyvän yksilön (FMNH P27393) New Mexicosta nimellä P. cyrtocristatus vuonna 1961. Se sisältää osittaisen kallon, jossa on lyhyt, pyöristetty harja, ja suuren osan kallon jälkeisestä luustosta lukuun ottamatta jalkoja, kaulaa ja osia hännästä. Erityisnimi on johdettu latinankielisistä sanoista curtus ”lyhennetty” ja cristatus ”harjattu”. Yksilö löydettiin joko Fruitlandin muodostuman yläosasta tai todennäköisemmin yläpuolisen Kirtlandin muodostuman pohjasta. Lajin levinneisyysalue laajeni vuonna 1979, kun David B. Weishampel ja James A. Jensen kuvasivat osittaisen kallon, jossa oli samanlainen harja (BYU 2467) Utahin Garfieldin piirikunnan (Garfield County, Utah) kampanjakautisesta Kaiparowitsin muodostumasta. Sittemmin Utahista on löydetty toinenkin kallo, jolla on lyhyt/pyöreä P. cyrtocristatus -harjan morfologia.
Laji
Parasaurolophus walkeri -kallonvalu, Natural History Museum, Lontoo.
Parasaurolophus cyrtocristatus kallonvalu, Field Museum of Natural History.
Tyyppilaji P. walkeri, Albertasta, tunnetaan yhdestä yksilöstä. Se eroaa P. tubicenista siinä, että sen harjanteessa on yksinkertaisemmat putket, ja P. cyrtocristatusista siinä, että sillä on pitkä, pyöristymätön harja ja pidempi ylävartalo kuin kyynärvarsi.
P. tubicen, New Mexicosta, tunnetaan ainakin kolmen yksilön jäännöksistä. Se on kookkain laji, jonka harjanteessa on monimutkaisemmat ilmakanavat kuin P. walkerilla ja pidempi ja suorempi harja kuin P. cyrtocristatuksella.
P. cyrtocristatus, New Mexicosta ja Utahista, tunnetaan kolmen mahdollisen yksilön perusteella. Se on pienin laji, jolla on lyhyt pyöreä harja. Sen pieni koko ja harjan muoto ovat saaneet useat tutkijat ehdottamaan, että se edustaa P. tubicenin, joka on peräisin suunnilleen samalta ajalta ja samasta muodostumasta New Mexicosta, nuoria yksilöitä tai naaraita. Kuten Thomas Williamson on todennut, P. cyrtocristatuksen tyyppiaineisto on noin 72 % P. tubicenin koosta, mikä on lähellä kokoa, jossa muiden lambeosauriinien on tulkittu alkavan osoittaa lopullista sukupuolidimorfismia harjanteissaan (~70 % aikuisen koosta). Tämä kanta on hylätty viimeaikaisissa lambeosaurineja koskevissa katsauksissa.
Paleobiologia
Paleoekologia
Gorgosaurus (vasemmalla) hyökkää Parasaurolophus cyrtocristatusin kimppuun.
Dinosauruspuiston muodostumasta peräisin oleva Parasaurolophus walkeri kuului monipuoliseen ja hyvin dokumentoituun esihistoriallisten eläinten faunaan, johon kuuluivat tunnetut dinosaurukset, kuten sarvipäiset Centrosaurus, Styracosaurus ja Chasmosaurus; lajitoverit Prosaurolophus, Gryposaurus, Corythosaurus ja Lambeosaurus, tyrannosaurus Gorgosaurus sekä panssaroidut Edmontonia ja Euoplocephalus. Se oli harvinainen osa tätä eläimistöä. Dinosaur Park -muodostuman tulkitaan olevan jokien ja tulva-alueiden matalareliefinen ympäristö, joka muuttui ajan mittaan soisemmaksi ja johon merelliset olosuhteet vaikuttivat, kun läntinen sisäinen meriväylä eteni länteen. Ilmasto oli lämpimämpi kuin nykyisessä Albertassa, jossa ei ollut pakkasta, mutta jossa oli kosteampia ja kuivempia vuodenaikoja. Havupuut olivat ilmeisesti vallitsevia latvuskasveja, ja aluskasvillisuudessa esiintyi saniaisia, saniaispuita ja angiospermejä.
Uuden Meksikon lajit jakoivat elinympäristönsä suurten sauropodien Alamosauruksen, ankkasyömäri Kritosauruksen, sarvipäisen Pentaceratopsin, panssaroidun Nodocephalosauruksen, Saurornitholesteksen ja tällä hetkellä nimeämättömän tyrannosauruksen kanssa. Kirtlandin muodostuman tulkitaan olevan jokien tulva-alueita, jotka ovat syntyneet läntisen sisäisen meriväylän vetäytymisen jälkeen. Havupuut olivat vallitsevia kasveja, ja chasmosauriiniset sarvipäiset dinosaurukset olivat ilmeisesti yleisempiä kuin hadrosauridit.
Ruokailu
Hadrosauridina Parasaurolophus oli suuri kaksi-/neljäjalkainen kasvinsyöjä, joka söi kasveja hienostuneella kallolla, joka mahdollisti pureskelua muistuttavan jauhamisliikkeen. Sen hampaat vaihtuivat jatkuvasti ja pakkautuivat hammasparistoihin, jotka sisälsivät satoja hampaita, joista vain suhteellisen pieni osa oli käytössä kerrallaan. Se käytti nokkaansa kasvimateriaalin sadonkorjuuseen, jota posken kaltainen elin piti leuassa kiinni. Ruokailu olisi tapahtunut maasta noin 4 metrin korkeuteen asti. Kuten Bob Bakker on todennut, lambeosauriinien nokka on kapeampi kuin hadrosauriinien, mikä viittaa siihen, että Parasaurolophus ja sen sukulaiset pystyivät ruokailemaan valikoivammin kuin niiden leveänokkaiset, harjaksettomat lajitoverit.
Kallonharja
Monia hypoteeseja on esitetty siitä, millaisia toimintoja Parasaurolophusin kallonharja suoritti, mutta suurin osa niistä on hylätty. Nykyään uskotaan, että sillä on saattanut olla useita tehtäviä: visuaalinen näyttö lajin ja sukupuolen tunnistamiseksi, äänen vahvistaminen viestintää varten ja lämmönsäätely. Ei ole selvää, mikä oli merkittävintä milloinkin harjan ja sen sisäisten nenäkäytävien evoluutiossa.
Lajien ja kasvuvaiheiden väliset erot
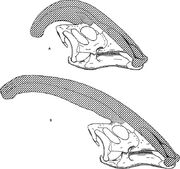
Diagrammi, jossa verrataan Parasaurolophus cyrtocristatusin (a) ja Parasaurolophus walkerin (b) nariaaliharjoja.
Kuten muidenkin lambeosauriinien kohdalla, Parasaurolophuksen kallonharjan uskotaan muuttuneen iän myötä ja olleen sukupuolisesti dimorfinen ominaisuus aikuisilla. James Hopson, yksi ensimmäisistä tutkijoista, joka kuvasi lambeosauriinien hartioita tällaisten erojen perusteella, esitti, että P. cyrtocristatus pienine hartioineen oli P. tubicen naarasmuoto. Thomas Williamson ehdotti, että se olisi nuorempi muoto. Kumpikaan hypoteesi ei tullut laajalti hyväksytyksi. Koska tiedossa on vain kuusi hyvää kalloa ja yksi nuori aivokotelo, lisäaineisto auttaa selvittämään näitä mahdollisia sukulaisuussuhteita. Williamson totesi, että joka tapauksessa nuorilla Parasaurolophuksilla oli todennäköisesti P. cyrtocristatuksen kaltaiset pienet, pyöreät harjanteet, jotka todennäköisesti kasvoivat nopeammin, kun yksilöt lähestyivät sukukypsyyttä. Aiemmin Lambeosaurukselle määritellyn ja nyt Parasaurolophukselle määritellyn nuoren aivokotelon äskettäinen uudelleentutkimus osoittaa, että nuorilla yksilöillä oli pieni putkimainen harja. Tässä näytteessä on säilynyt pieni ylöspäin suuntautuva otsaluun levennys, joka oli samankaltainen mutta pienempi kuin aikuisilla yksilöillä; aikuisilla yksilöillä otsaluut muodostivat alustan, joka kannatteli harjanteen pohjaa. Tämä näyte osoittaa myös, että Parasaurolophuksen harjan kasvu ja nuorten yksilöiden kasvoprofiili poikkesivat Corythosaurus-Hypacrosaurus-Lambeosaurus-mallista osittain siksi, että Parasaurolophuksen harjalta puuttuu ohut luinen ”coxcomb”, joka muodostaa kolmen muun lambeosauruksen harjan yläosan.
Hylätyt toimintahypoteesit
Monissa varhaisissa ehdotuksissa keskityttiin vesielämäntapaan liittyviin sopeutumisiin sen hypoteesin mukaisesti, jonka mukaan hadrosauridit olivat sammakkoeläimiä, mikä oli yleinen ajatuslinja 1960-luvulle asti. Niinpä Alfred Sherwood Romer ehdotti, että se toimi snorkkelina, Martin Wilfarth, että se oli kiinnitys liikkuvalle sorkkaraudalle, jota käytettiin hengitysputkena tai ruoan keräämiseen, Charles M. Sternberg, että se toimi ilmansulkuna pitääkseen veden poissa keuhkoista, ja Ned Colbert, että se toimi ilmasäiliönä pidempiaikaista veden alla oleskelua varten.
Muut ehdotukset olivat luonteeltaan fyysisempiä. Kuten edellä mainittiin, William Parks ehdotti, että se oli liitetty nikamiin nivelsiteillä tai lihaksilla ja auttoi pään liikuttamisessa ja tukemisessa. Othenio Abel ehdotti, että sitä käytettiin aseena saman lajin jäsenten välisessä taistelussa, ja Andrew Milner ehdotti, että sitä voitiin käyttää lehtien suojana, kuten kasuaarin kypärävaakunaa (nimeltään ”casque”). Toisissa ehdotuksissa tärkeimmäksi tehtäväksi asetettiin erikoistuneiden elinten asentaminen. Halszka Osmólska ehdotti, että siinä sijaitsivat suolarauhaset, ja John Ostrom ehdotti, että siinä sijaitsivat laajemmat alueet hajukudokselle ja lambeosauriinien paljon paremmalle hajuaistille, jolla ei ollut ilmeisiä puolustusvalmiuksia. Eräs epätavallinen, kreationisti Duane Gishin esittämä ehdotus on, että harjanteessa oli kemiallisia rauhasia, joiden avulla se pystyi heittämään kemiallisia ”tulisuihkuja” vihollisia kohti, samaan tapaan kuin nykyajan pommikovakuoriainen.
Useimmat näistä hypoteeseista on diskreditoitu tai hylätty. Esimerkiksi harjanteen päässä ei ole reikää snorklaustoimintoa varten. Ei ole lihasarpia sorkkarauhasta varten, ja on kyseenalaista, että eläin, jolla on nokka, tarvitsisi sellaista. Ehdotettuna ilmalukkona se ei olisi pitänyt vettä ulkona. Ehdotettu ilmasäiliö olisi ollut riittämätön Parasaurolophuksen kokoiselle eläimelle. Muilla hadrosaurideilla oli suuret päät ilman, että ne olisivat tarvinneet suuria onttoja harjanteita, jotka olisivat toimineet kiinnityskohtina tukiville nivelsiteille. Mikään ehdotuksista ei myöskään selitä, miksi harjanteella on tällainen muoto, miksi muilla lambeosaurinoilla pitäisi olla harjanteet, jotka näyttävät paljon erilaisilta mutta joilla on samanlainen tehtävä, miten harjanteettomat tai kiinteäharjanteiset hadrosauridit tulivat toimeen ilman tällaisia ominaisuuksia tai miksi joillakin hadrosaurideilla oli kiinteät harjanteet. Nämä seikat vaikuttavat erityisesti hypoteeseihin, jotka perustuvat eläimessä jo olemassa olevien järjestelmien kykyjen lisäämiseen, kuten suolarauhashypoteesiin ja hajuhypoteesiin, ja viittaavat siihen, että nämä eivät olleet harjanteen ensisijaisia tehtäviä. Lisäksi lambeosauriinien nenäontelosta tehdyt työt osoittavat, että hajuhermot ja vastaavat aistikudokset olivat suurelta osin harjanteeseen kuuluvan nenäkäytävän osan ulkopuolella, joten harjanteen laajentumisella ei ollut juurikaan tekemistä hajuaistin kanssa.
Sosiaaliset toiminnot
Life restoration of Parasaurolophus walkeri with hypothetical skin frill and stripes on its crest.
Sosiaaliset ja fysiologiset toiminnot ovat sen sijaan saaneet enemmän kannatusta harjanteen toiminto(i)na keskittyen visuaaliseen ja auditiiviseen tunnistamiseen ja kommunikointiin. Suurena esineenä harjalla on selkeä arvo visuaalisena signaalina, ja se erottaa tämän eläimen aikalaisistaan. Hadrosauridien silmäkuoppien suuri koko ja skleroottisten renkaiden esiintyminen silmissä viittaavat terävään näkökykyyn ja päiväsaikaan, mikä on osoitus siitä, että näkö oli näille eläimille tärkeää. Jos, kuten tavallisesti kuvataan, nahkahapsut ulottuivat harjanteelta kaulalle tai selkään, ehdotettu näkönäytös olisi ollut paljon näyttävämpi. Kuten muiden lambeosaurusten kallot viittaavat, Parasaurolophuksen harja mahdollisti todennäköisesti sekä lajintunnistuksen (kuten sen erottamisen Corythosauruksesta tai Lambeosauruksesta) että urosten ja naaraiden välisen määrittelyn muodon ja koon perusteella.
Ohjaustoiminta
Tietokonemalli P. tubicen harjanteen nenäkäytävistä.
Harjanteen ulkoinen ulkomuoto ei kuitenkaan vastaa nenäkäytävien monimutkaista sisäistä anatomiaa, mikä viittaa johonkin muuhun toimintoon, joka selittää sisätilan käytön. Carl Wiman ehdotti ensimmäisenä vuonna 1931, että nenäkäytävät palvelivat kuuloaistiin liittyvää signaalitehtävää, kuten käyrätorvi; Hopson ja David B. Weishampel palasivat tähän ajatukseen 1970- ja 1980-luvuilla. Hopson totesi, että on olemassa anatomisia todisteita siitä, että hadrosaurideilla oli vahva kuulo. Ainakin yksi esimerkki, sukulaiseläimessä Corythosauruksessa, osoittaa, että matelijoiden korvaluu (stapes) on paikallaan, mikä yhdistettynä tärykalvolle varattuun suureen tilaan viittaa herkkään välikorvaan. Lisäksi hadrosauridin lagena on krokotiilin kaltainen pitkänomainen, mikä viittaa siihen, että sisäkorvan kuulo-osa oli hyvin kehittynyt. Weishampel esitti, että P. walkeri pystyi tuottamaan taajuuksia 48-240 Hz ja P. cyrtocristatus (tulkittuna nuorena harjumuotona) 75-375 Hz. Perustuen hadrosauridien sisäkorvien samankaltaisuuteen krokotiilien sisäkorvien kanssa hän ehdotti myös, että aikuiset hadrosauridit olivat herkkiä korkeille taajuuksille, jollaisia niiden jälkeläiset saattavat tuottaa. Weishampelin mukaan tämä on sopusoinnussa vanhempien ja jälkeläisten kommunikoinnin kanssa.
Tietokonemallinnus hyvin säilyneestä P. tubicen -yksilöstä, jolla on monimutkaisemmat hengityskanavat kuin P. walkeri -yksilöllä, on mahdollistanut sen mahdollisen äänen rekonstruoinnin, jonka sen harja tuotti. Pääreitti resonoi noin 30 Hz:n taajuudella, mutta monimutkainen sinuksen anatomia aiheuttaa äänessä piikkejä ja laaksoja.
Jäähdytystoiminto
Harjun suuri pinta-ala ja verisuonitus viittaavat myös lämmönsäätelyyn. P.E. Wheeler ehdotti tätä käyttöä ensimmäisen kerran vuonna 1978 keinona pitää aivot viileinä. Myös Teresa Maryańska ja Osmólska ehdottivat lämmönsäätelyä suunnilleen samaan aikaan, ja Sullivan ja Williamson kiinnostuivat lisää. David Evansin vuonna 2006 esittämä keskustelu lambeosaurusten harjanteiden toiminnoista puolsi ajatusta, ainakin harjanteiden laajenemisen evoluution alkutekijänä.
Mediassa
Parasaurolophus on hyvin suosittu dinosaurus. Se oli elokuvassa Disneyn Dinosaurus lauman jäsenenä. Se esiintyi myös useita kertoja kuuluisassa dokumentissa Clash of the Dinosaurs. Se esiintyi myös suositussa TV-sarjassa Prehistoric Park, jossa siitä tuli suurten lihansyöjien Deinosuchuksen ja Albertosauruksen saalis. Se esiintyi muutaman kerran Jurassic Park -elokuvissa: ensimmäisessä elokuvassa se oli lauman jäsen, toisessa elokuvassa se jäi metsästäjien vangiksi ja kolmannessa elokuvassa se juoksi yhdessä Corythosauruksen kanssa. ja hahmona Dweeb elokuvassa We’re Back! A Dinosaur’s Story”, on itse Parasaurolophus.
Parasaurolophus esiintyy myös Turok-sarjassa, rauhallisena kasvinsyöjänä, joka ei normaalisti ole haitallinen, mutta voi olla aggressiivinen, jos häntä ärsytetään vakavasti.
Power Rangers -sarjassa on myös Parasaurolophus zord: Dino Thunder ja Power Rangers: Dino Charge.
Parasaurolophus esiintyy lyhyesti Disneyn Pixarin elokuvan The Good Dinosaur alussa.
See Also
Parasaurolophus/Galleria
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, sivu 229. Julkaistu: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, luku: Parasaurolophus, sivut 678-684. Julkaistu: 1997, McFarland & Co, Jefferson, Pohjois-Carolina. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull ja Wright, Hadrosaurian Dinosaurs of North America, s. 209-213.
- Brett-Surman, Michael K. ja Wagner, Jonathan R. Carpenter, Kenneth (toim.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, luku: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, sivut 135-169. Julkaistu, 2006, Indiana University Press, Bloomington ja Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 5.5 Horner, John R., Weishampel, David B. ja Forster, Catherine A, Weishampel, David B., Osmólska, Halszka ja Dodson, Peter (toim.) The Dinosauria, 2. painos, luku: Hadrosauridae, sivut 438-463. Julkaistu: 2004, University of California Press, Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, volume 13, pages 1-32. Julkaistu: 1922, University of Toronto Studies, Geology Series.
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. ja Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, sarjasta New Mexicon luonnonhistoriallisen museon ja luonnontieteellisen tiedelehden tiedotuslehti (New Mexico Museum of Natural History and Science Bulletin), julkaisusarja, 15, sivut 1-52. Julkaistu: 1999, New Mexico Museum of Natural History and Science, Albuquequeque, New Mexico.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, Journal of Paleontology, Volume 35, 3rd issue, on pages 575-577.
- Gilmore, Charles W., On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, volume 38, issue 43, pages 29-48, Parks. Julkaistu: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. ja Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; ja Dodson, Peter (toim.) The Dinosauria, 1. painos, Chapter: Hadrosauridae, sivut 534-561. Julkaistu: 1990, University of California Press, Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., ja Reisz, Robert R. 2007. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta, from the Journal of Vertebrate Paleontology, vol. 27 issue 2, on pages 373-393.
- Godefroit, Pascal, Shuqin Zan; ja Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China, from the Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, pages 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Julkaisija New Mexico Museum of Natural History and Science, in Albuquequeque, New Mexico. Sivut 191-213.
- Liddell, Henry George ja Robert Scott, 1980. A Greek-English Lexicon (lyhennetty painos) Julkaistu: Oxford University Press Yhdistyneessä kuningaskunnassa. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. ja Evans, David C., Currie, Phillip J. ja Koppelhus, Eva (toim.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, luku: Ornithischian Dinosaurs. 2005, Julkaistu: Indiana University Press, Bloomington. Sivut 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; ja Dupuis, Kevin, 2007. A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus. Journal of Vertebrate Paleontology, vol. 27, issue 3 pages 642-650.
- Currie, Phillip J; and Russell, Dale A. ”Geographic and stratigraphic distribution of dinosaur remains” in Dinosaur Provincial Park, s. 553.
- Weishampel, David B. (1990). ”Dinosaurusten levinneisyys”, teoksessa The Dinosauria (1.), s. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; ja Noto, Christopher R. (2004). ”Dinosaurusten levinneisyys”, teoksessa The Dinosauria (2.), s. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, julkaisusta Nova Acta Regia Societas Scientarum Upsaliensis, series 4, vol. 7, issue 5. (saksa). Sivut 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, julkaissut Cassell Ltd. 1979, painos 5, Lontoossa. ISBN 0-304-52257-0 Sivu 883.
- Weishampel, David B. ja Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) Utahista, julkaisusta Journal of Paleontology, vol. 53, issue 6, pages 1422-1427.
- Eberth, David A. 2005. ”Geologia”, teoksessa Dinosaur Provincial Park, s. 54-82.
- Braman, Dennis R. ja Koppelhus, Eva B. 2005. ”Campanian palynomorphs”, teoksessa Dinosaur Provincial Park, s. 101-130.
- Russell, Dale A. An Odyssey in Time: Pohjois-Amerikan dinosaurukset, 1989. Kustantaja: NorthWord Press, Minocqua, Wisconsin. ISBN 1-55971-038-1 Sivut 160-164.
- Bakker, Robert T. 1986. Dinosaurusten harhaopit: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, julkaisija William Morrow, New York. ISBN 0-8217-2859-8 Sivu 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: Original and Compelling Insight into Life in the Dinosaur Kingdom, luku: Hadrosaurids II. 1985. Julkaisija: Crescent Books, New York. Sivut 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs, julkaisusta Journal of Paleobiology, vol. 32, issue 1, Pages 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs , from the Journal of Paleobiology, volume one, issue one, Pages 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology , University of Chicago Pressistä, sivulla 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, julkaisusta Journal of Orion (München), vol. 2. s. 525-532 (saksankielinen).
- Sternberg, Charles M. 1935. Belly River Series of the Belly River Series of the Upper Cretaceous from the Journal of the Canada Department of Mines Bulletin (Geological Series), volume 77, issue 52, on pages 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, julkaissut vuonna 1945 American Museum of Natural History, Man and Nature Publications, 14, New York. Sivu 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas from the Journal of Jarbuch Naturwissenschaften, volume 12, issue 36, on pages 709-716. (saksa) 1924.
- Osmólska, Halszka, 1979. Nasaaliset suolarauhaset dinosauruksilla, lehdestä Acta Palaeontologica Polonica, vol. 24, sivut 205-215.
- 36.0 36.1 Ostrom, John H., 1962. The cranial crests of hadrosaurian dinosaurs, alkaen Journal of Postilla, vol. 62, sivut 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, kustantaja Master Books, Green Forest. ISBN 0-89051-165-9 Sivu 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), julkaisusta Journal of Paleobiology, vol. 7, issue 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, alkaen Journal of Paleontology, vol. 55, issue 5, pp. 1046-1057.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur, Sandia National Laboratories (1997-12-05). Haettu 20. tammikuuta 2009.
- Diegert, Carl F. ja Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen alkaen Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. ja Williamson, Thomas E., 1996. A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications (Uusi Parasaurolophuksen kallo (pitkävartinen muoto) Uudesta Meksikosta: ulkoiset ja sisäiset (CT-kuvaukset) piirteet ja niiden toiminnalliset merkitykset), julkaisusta Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborate CNS cooling structure in large dinosaurs Journal of Nature, vol. 275, on pp. 441-443.
- Maryańska, Teresa ja Osmólska, Halszka, vuonna 1979. Aspects of hadrosaurian cranial anatomy, Journal of Lethaia, vol. 12, on pp. 265-273.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur; from Sandia National Laboratories.
- Restorations of P. walkeri and P. cyrtocristatus, to the same scale, by Scott Hartman; at Skeletal Drawing.com.
- Parasaurolophus page on Enchanted Learning.com