

Parasaurolophus (ejtsd: /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, általában még /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; jelentése “közeli címeres gyík”, a Saurolophusra utalva) egy ornithopoda dinoszaurusz nemzetség a mai Észak-Amerika területének késő kréta időszakából, körülbelül 76-73 millió évvel ezelőttről. Növényevő állat volt, amely két- és négylábúként is közlekedett. Három faját ismerik: P. walkeri (a típusfaj), P. tubicen, és a rövidhátú P. cyrtocristatus. A maradványok a kanadai Albertából, valamint az egyesült államokbeli Új-Mexikóból és Utahból ismertek. Először 1922-ben írta le William Parks egy koponya és részleges csontváz alapján Albertában.
A Parasaurolophus egy hadrosauridafaj, a bizarr fejdíszeiről ismert kréta kori dinoszauruszok változatos családjának tagja. Ez a nemzetség nagy, kidolgozott koponyacsúcsáról ismert, amely a legnagyobb méretben egy hosszú, ívelt csövet alkot, amely felfelé és hátrafelé nyúlik ki a koponyából. A Kínából származó Charonosaurusnak, amely a legközelebbi rokona lehetett, hasonló koponyája és valószínűleg hasonló fejdísze volt. A tudósok sokat vitatkoztak a címerről; a konszenzus szerint a fő funkciók közé tartozott a faj és a nem vizuális felismerése, az akusztikus rezonancia és a hőszabályozás. A ritkább kacsacsőrűek közé tartozik, csak néhány jó példányról ismert.
A Parasaurolophus a legismertebb dinoszauruszok közé tartozik, és gyakran ábrázolják a dinoszauruszokkal kapcsolatos televíziós műsorokban, filmekben, könyvekben, árucikkekben és más médiumokban. Alice Perry
leírás

A Parasaurolophus walkeri mérete az emberéhez képest.
Mint a legtöbb dinoszaurusz esetében, a Parasaurolophus csontváza is hiányosan ismert. A P. walkeri típuspéldányának hosszát 9,5 méterre becsülik. A koponyája a címerrel együtt körülbelül 1,6 méter hosszú, míg a P. tubicen típuskoponyája több mint 2,0 méter hosszú, ami nagyobb állatra utal. Tömegét 2,5 tonnára becsülik. Egyetlen ismert mellső végtagja hadrosauridához képest viszonylag rövid volt, rövid, de széles lapockával. A combcsont a P. walkeri esetében 103 centiméter hosszú, és más hadrosauridákhoz képest hosszúságához képest robusztus. A felkar és a medencecsontok szintén erősen felépítettek.
A többi hadrosauridához hasonlóan képes volt két vagy négy lábon járni. Valószínűleg inkább négy lábon kereste a táplálékot, de két lábon futott. A csigolyák idegcsigolyái magasak voltak, ahogy az a lambeosaurináknál szokásos volt; a legmagasabbak a csípő felett voltak, és növelték a hát magasságát. A P. walkeri bőrének lenyomatai ismertek, amelyek egyenletes, gumószerű pikkelyeket mutatnak, de nagyobb struktúrákat nem.
Parasaurolophus walkeri a pikkelyezés részleteivel.
A legfeltűnőbb jellegzetesség a koponyacsúcs volt, amely a fej hátsó részéből állt ki, és a premaxillából és az orrcsontokból állt. A P. walkeri típuspéldányon az idegtüskékben van egy bevágás, közel ahhoz a helyhez, ahol a gerinc a hátuljához érne, de ez lehet, hogy csak erre az egyedre jellemző patológia. William Parks, aki a nemzetség nevét adta, azt feltételezte, hogy egy szalag fut a gerincoszloptól a bevágásig, hogy megtámassza a fejet. Bár ez az elképzelés valószínűtlennek tűnik, a Parasaurolophus néha a gerinctől a nyakig tartó bőrlebennyel áll helyre.
A gerinc üreges volt, mindkét orrlyukból különálló csövek vezettek a gerinc végéig, mielőtt megfordították volna az irányt, és visszamentek volna a gerincen lefelé a koponyába. A csövek a P. walkeri esetében voltak a legegyszerűbbek, a P. tubicen esetében pedig összetettebbek, ahol egyes csövek vakok voltak, mások pedig találkoztak és szétváltak. Míg a P. walkeri és a P. tubicen hosszú, csak enyhén görbült címerrel rendelkezett, addig a P. cyrtocristatus rövid, inkább körkörös profilú címerrel.
Klasszifikáció
A Parasaurolophusról – ahogy a neve is mutatja – kezdetben azt gondolták, hogy a Saurolophusszal áll közeli rokonságban a felületesen hasonló címer miatt. Hamarosan azonban újraértékelték, hogy a hadroszauruszok lambeoszaurusz alcsaládjába tartozik – a Saurolophus egy hadroszaurusz. Általában a lambeosaurusok különálló ágaként értelmezik, amely elkülönül a sisakos Corythosaurustól, a Hypacrosaurustól és a Lambeosaurustól. Legközelebbi ismert rokonának a Charonosaurus tűnik, egy hasonló koponyával (de még nem teljes címerrel) rendelkező lambeosaurus az északkelet-kínai Amur-vidékről, és a kettő a Parasaurolophini kládot alkothatja. A P. cyrtocristatus a rövid, kerekebb címerével a három ismert Parasaurolophus faj közül a legbazálisabb lehet, vagy a P. tubicen szubadult vagy nőstény példányait képviselheti.
Történet
Felfedezés és elnevezés
A Parasaurolophus walkeri hiányos típuspéldányának öntvénye Warszawában.
A Parasaurolophus név jelentése “közeli címeres gyík”, a görög para/παρα “mellett” vagy “közel”, saurus/σαυρος “gyík” és lophos/λοφος “címer” szavakból származik. Az elnevezés alapja a ROM 768-as számú koponya és részleges csontváz, amelyből hiányzik a farok nagy része és a térd alatti hátsó lábak, és amelyet a Torontói Egyetem kutatócsoportja talált 1920-ban a kanadai Alberta államban, a Red Deer folyó menti Sand Creek közelében. Ezeket a kőzeteket ma a Campanian korú felső kréta korú Dinosaur Park Formation néven ismerjük. William Parks a példányt Sir Byron Edmund Walker, a Királyi Ontariói Múzeum kuratóriumának elnöke tiszteletére nevezte el P. walkeri-nek. A Parasaurolophus maradványai ritkák Albertában, csak egy másik részleges koponya származik (valószínűleg) a Dinosaur Park Formációból, és három Dinosaur Park példányból hiányzik a koponya, amelyek valószínűleg a nemzetséghez tartoznak. Egyes faunalistákban említést tesznek a P. walkeri lehetséges anyagáról a montanai Hell Creek Formációban, egy késő maastrichti korú kőzetegységben. Ezt az előfordulást Sullivan és Williamson a nemzetségről szóló 1999-es áttekintésükben nem említik, és máshol sem foglalkoztak vele részletesebben.
1921-ben Charles H. Sternberg egy részleges koponyát (PMU.R1250) talált az Új-Mexikóban, San Juan megyében található, ma kissé fiatalabb Kirtland Formációból. Ezt a példányt a svédországi Uppsalába küldték, ahol Carl Wiman 1931-ben egy második fajként, P. tubicen néven írta le. A faj specifikus epitheton a latin tǔbǐcěn “trombitás” szóból származik. Egy második, majdnem teljes P. tubicen koponyát (NMMNH P-25100) 1995-ben találtak Új-Mexikóban. E koponya komputertomográfiájának felhasználásával Robert Sullivan és Thomas Williamson 1999-ben monografikus feldolgozást adott a nemzetségnek, kitérve az anatómiai és taxonómiai szempontokra, valamint a gerinc funkcióira. Williamson később egy független áttekintést publikált a maradványokról, amelyben nem értett egyet a rendszertani következtetésekkel.
John Ostrom 1961-ben egy másik jó példányt (FMNH P27393) Új-Mexikóból P. cyrtocristatus néven írt le. Tartalmaz egy részleges koponyát rövid, lekerekített címerrel, valamint a koponya utáni csontváz nagy részét, kivéve a lábakat, a nyakat és a farok egyes részeit. Különleges neve a latin curtus “megrövidült” és cristatus “csúcsos” szóból származik. A példányt vagy a Fruitland Formáció tetején, vagy – ami valószínűbb – a felette lévő Kirtland Formáció alján találták. A faj elterjedési területe 1979-ben bővült, amikor David B. Weishampel és James A. Jensen leírtak egy hasonló címerrel rendelkező részleges koponyát (BYU 2467) a Utah állambeli Garfield megyében található kampán korú Kaiparowits Formációból. Azóta Utahban egy másik koponyát is találtak a P. cyrtocristatus rövid/kerek címerének morfológiájával.
Species
Parasaurolophus walkeri koponyaöntvény, Natural History Museum, London.
Parasaurolophus cyrtocristatus koponyaöntvény, Field Museum of Natural History.
A típusfaj P. walkeri, Albertából, egyetlen példányról ismert. A P. tubicentől abban különbözik, hogy egyszerűbb csövek vannak a címerében, a P. cyrtocristatustól pedig abban, hogy a címer hosszú, nem lekerekített és a felkar hosszabb, mint az alkar.
A P. tubicen Új-Mexikóból legalább három egyed maradványaiból ismert. Ez a legnagyobb faj, a P. walkerihez képest bonyolultabb légutakkal a címerében, és hosszabb, egyenesebb címerrel, mint a P. cyrtocristatus.
A P. cyrtocristatus, Új-Mexikóból és Utahból, három lehetséges példányból ismert. Ez a legkisebb faj, rövid, lekerekített címerrel. Kis mérete és a címer formája több tudóst arra késztetett, hogy azt feltételezzék, hogy a P. tubicen fiatal egyedeit vagy nőstényeit képviseli, amely nagyjából ugyanabból az időből és ugyanabból az új-mexikói formációból származik. Ahogy Thomas Williamson megjegyezte, a P. cyrtocristatus típusanyaga a P. tubicen méretének kb. 72%-a, ami közel van ahhoz a mérethez, amelynél más lambeosaurine-oknál az értelmezések szerint elkezdődik a határozott nemi dimorfizmus a címerben (a felnőtt méret ~70%-a). Ezt az álláspontot a lambeosaurinák legújabb áttekintései elvetették.
Paleobiológia
Paleoökológia
Gorgosaurus (balra) megtámad egy Parasaurolophus cyrtocristatus-t.
A Dinosaur Park Formációból származó Parasaurolophus walkeri az őskori állatok változatos és jól dokumentált faunájának tagja volt, köztük olyan jól ismert dinoszauruszoké, mint a szarvú Centrosaurus, a Styracosaurus és a Chasmosaurus; a Prosaurolophus, a Gryposaurus, a Corythosaurus és a Lambeosaurus; a tyrannosaurid Gorgosaurus; valamint a páncélos Edmontonia és Euoplocephalus. Ennek a faunának ritka alkotóeleme volt. A Dinosaur Park Formáció értelmezése szerint a folyók és árterek alacsony domborzatú környezetében alakult ki, amely az idő múlásával egyre mocsarasabbá vált, és tengeri körülmények befolyásolták, ahogy a nyugati Belső-tengeri út nyugat felé haladt. Az éghajlat melegebb volt, mint a mai Alberta területén, fagyok nélkül, de csapadékosabb és szárazabb évszakokkal. A jelek szerint a tűlevelűek voltak a domináns lombkoronanövények, az aljnövényzetet páfrányok, fapáfrányok és angiospermák alkották.
Az új-mexikói fajok a nagy szauropoda Alamosaurus, a kacsacsőrű Kritosaurus, a szarvas Pentaceratops, a páncélos Nodocephalosaurus, a Saurornitholestes és a jelenleg meg nem nevezett tyrannosauridák környezetében éltek. A Kirtland-képződményt a Nyugati Belső-tengerszoros visszahúzódása után megjelenő folyami árterekként értelmezik. A tűlevelűek voltak a domináns növények, és a chasmosaurus szarvú dinoszauruszok nyilvánvalóan gyakoribbak voltak, mint a hadrosauridák.
Táplálkozás
A Parasaurolophus hadrosauridaként egy nagy két-/négylábú növényevő volt, amely növényeket fogyasztott, kifinomult koponyával, amely lehetővé tette a rágással analóg őrlő mozgást. Fogai folyamatosan cserélődtek, és több száz fogat tartalmazó fogakba tömörültek, amelyek közül mindig csak viszonylag kevés volt használatban. A csőrét használta a növényi anyag learatására, amelyet egy pofaszerű szerv tartott az állkapcsában. A táplálkozás a földről kb. 4 méteres magasságig történhetett. Ahogy Bob Bakker megjegyezte, a lambeosaurinák csőre keskenyebb, mint a hadroszaurináké, ami arra utal, hogy a Parasaurolophus és rokonai szelektívebben táplálkozhattak, mint széles csőrű, csőr nélküli társaik.
Koponyacsőr
Már számos hipotézis született arra vonatkozóan, hogy a Parasaurolophus koponyacsőrének milyen funkciói voltak, de a legtöbbet hiteltelenítették. Ma már úgy vélik, hogy több funkciója is lehetett: vizuális megjelenítés a faj és a nem azonosítására, hangerősítés a kommunikációhoz és hőszabályozás. Nem világos, hogy a címer és a belső orrjáratok evolúciójában melyik volt a legjelentősebb, melyik időpontban.
A fajok és növekedési szakaszok közötti különbségek
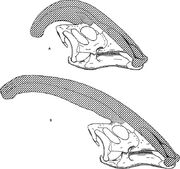
A Parasaurolophus cyrtocristatus (a) és a Parasaurolophus walkeri (b) náriális címerét összehasonlító ábra.
A többi lambeosaurinához hasonlóan úgy vélik, hogy a Parasaurolophus koponyacsúcsa az életkorral változott, és a felnőtteknél nemi dimorf jellegzetesség volt. James Hopson, az egyik első kutató, aki a lambeosaurusok címerét ilyen megkülönböztetéssel írta le, azt javasolta, hogy a kis címerrel rendelkező P. cyrtocristatus a P. tubicen nőstény formája volt. Thomas Williamson szerint ez volt a juvenilis forma. Egyik hipotézis sem vált széles körben elfogadottá. Mivel csak hat jó koponya és egy juvenilis agykoponya ismert, további anyag segít tisztázni ezeket a lehetséges kapcsolatokat. Williamson megjegyezte, hogy mindenesetre a fiatalkorú Parasaurolophusnak valószínűleg a P. cyrtocristatushoz hasonlóan kicsi, lekerekített gerincük volt, amely valószínűleg gyorsabban nőtt, ahogy az egyedek közeledtek az ivarérettséghez. Egy korábban a Lambeosaurushoz, most pedig a Parasaurolophushoz rendelt fiatalkori agykoponya közelmúltbeli újratanulmányozása bizonyítékot szolgáltat arra, hogy a fiatalkorúaknál jelen volt egy kis csöves címer. Ez a példány megőrizte a homlokcsontok kis felfelé szélesedését, amely hasonló, de kisebb volt, mint a kifejlett példányoknál megfigyelhető; a felnőtteknél a homlokcsontok egy platformot alkottak, amely a homlokcsontok alapját támasztotta alá. Ez a példány azt is jelzi, hogy a Parasaurolophus gerincének növekedése és a fiatal egyedek arcprofilja eltért a Corythosaurus-Hypacrosaurus-Lambeosaurus modelltől, részben azért, mert a Parasaurolophus gerincéből hiányzik a vékony csontos “coxcomb”, amely a másik három lambeosaurus gerincének felső részét alkotja.
A funkcióval kapcsolatos elvetett hipotézisek
Sok korai felvetés a vízi életmódhoz való alkalmazkodásra összpontosított, azt a hipotézist követve, hogy a hadrosauridák kétéltűek voltak, ami az 1960-as évekig elterjedt gondolatmenet volt. Így Alfred Sherwood Romer azt javasolta, hogy légzőcsőként szolgált, Martin Wilfarth szerint egy mozgó ormányhoz való rögzítés volt, amelyet légzőcsőként vagy táplálékgyűjtéshez használtak, Charles M. Sternberg szerint légcsapdaként szolgált, hogy távol tartsa a vizet a tüdőtől, Ned Colbert szerint pedig légtartályként szolgált a hosszabb víz alatti tartózkodáshoz.
Más javaslatok inkább fizikai jellegűek voltak. Mint már említettük, William Parks azt javasolta, hogy szalagokkal vagy izmokkal kapcsolódott a csigolyákhoz, és segített a fej mozgatásában és alátámasztásában. Othenio Abel azt javasolta, hogy fegyverként használták az azonos faj tagjai közötti harcban, Andrew Milner pedig azt javasolta, hogy lombterelőként használhatták, mint a kazuár sisakpajzsát (amelyet “casque”-nak neveznek). Más javaslatok szerint a fő funkciót a speciális szervek elhelyezése jelentette. Halszka Osmólska felvetette, hogy sómirigyeknek adott otthont, John Ostrom pedig azt, hogy a szaglószövetnek és a lambeoszauruszok sokkal jobb szaglásának kibővített területeinek adott otthont, amelyek nem rendelkeztek nyilvánvaló védekező képességekkel. Egy szokatlan felvetés, amelyet a kreacionista Duane Gish tett, az, hogy a címer kémiai mirigyeknek adott otthont, amelyek lehetővé tették számára, hogy kémiai “tűzsugarakat” dobjon az ellenségre, hasonlóan a mai bombardier bogárhoz.
A legtöbb ilyen hipotézist diszkreditálták vagy elvetették. Például a címer végén nincs olyan lyuk, amely sznorkelező funkciót látott volna el. Nincsenek izomhegek az ormányoscsőrhöz, és kétes, hogy egy csőrrel rendelkező állatnak szüksége lenne rá. Mint javasolt légzsilip, nem tartotta volna távol a vizet. A javasolt légtartály nem lett volna elegendő egy Parasaurolophus méretű állat számára. Más hadrosauridáknak nagy fejük volt, anélkül, hogy nagy üreges csőrcsúcsokra lett volna szükségük, amelyek a tartószalagok rögzítési pontjaiként szolgáltak volna. Emellett a javaslatok egyike sem magyarázza meg, hogy miért van ilyen alakú a gerinc, hogy más lambeoszauruszoknak miért kellene másképp kinéző, de hasonló funkciót betöltő gerincekkel rendelkezniük, hogy a gerinc nélküli vagy tömör gerincű hadroszauruszok hogyan boldogultak ilyen képességek nélkül, vagy hogy egyes hadroszauruszoknak miért volt tömör gerincük. Ezek a megfontolások különösen hatással vannak az állatban már meglévő rendszerek képességeinek növelésén alapuló hipotézisekre, mint például a sómirigy és a szaglás hipotézisére, és arra utalnak, hogy ezek nem a címer elsődleges funkciói voltak. Ezenkívül a lambeosaurusok orrüregével kapcsolatos munkák azt mutatják, hogy a szaglóidegek és a megfelelő érzékszövetek nagyrészt az orrjáratoknak a gerincben lévő részén kívül voltak, így a gerinc kiterjedésének kevés köze volt a szagláshoz.
Társadalmi funkciók
Life restoration of Parasaurolophus walkeri with hypothetical skin frill and stripes on its crest.
Ehelyett a társadalmi és fiziológiai funkciókat támogatták inkább, mint a címer funkció(i)t, a vizuális és auditív azonosításra és kommunikációra összpontosítva. Nagyméretű tárgyként a címer egyértelmű vizuális jelzésértékkel bír, és megkülönbözteti ezt az állatot kortársaitól. A hadrosauridák szemgödreinek nagy mérete és a szklerotikus gyűrűk jelenléte a szemeken éles látásra és nappali szokásokra utal, ami azt bizonyítja, hogy a látás fontos volt ezeknek az állatoknak. Ha – ahogyan azt általában ábrázolják – egy bőrfodor húzódott a gerincről a nyakra vagy a hátra, akkor a javasolt vizuális megjelenítés sokkal látványosabb lett volna. Ahogy azt más lambeosaurus koponyák is sugallják, a Parasaurolophus címer valószínűleg lehetővé tette mind a faj azonosítását (például a Corythosaurustól vagy Lambeosaurustól való elkülönítését), mind a hímek és nőstények közötti meghatározást az alak és a méret alapján.
Hangfunkció
A P. tubicen orrjáratainak számítógépes modellje a gerincben.
A gerinc külső megjelenése azonban nem felel meg az orrjáratok bonyolult belső anatómiájának, ami arra utal, hogy a belső tér használatáért más funkció felelős. Carl Wiman volt az első, aki 1931-ben azt javasolta, hogy a járatok hallásjelző funkciót töltenek be, mint egy kürt; Hopson és David B. Weishampel az 1970-es és 1980-as években újra felvetette ezt az elképzelést. Hopson megállapította, hogy anatómiai bizonyítékok vannak arra, hogy a hadrosauridák erős hallással rendelkeztek. Legalább egy példa van arra, hogy a rokon Corythosauruson egy karcsú kengyel (hüllőfülcsont) van a helyén, ami a dobhártya számára kialakított nagy térrel együtt érzékeny középfülre utal. Továbbá a hadrosauridák lagénája a krokodilokéhoz hasonlóan hosszúkás, ami arra utal, hogy a belső fül hallószervi része jól fejlett volt. Weishampel szerint a P. walkeri 48 és 240 Hz közötti, a P. cyrtocristatus (amelyet egy ifjúkori címeres formaként értelmeztek) pedig 75 és 375 Hz közötti frekvenciák előállítására volt képes. A hadrosauridák belső fülének a krokodilokéhoz való hasonlósága alapján azt is javasolta, hogy a kifejlett hadrosauridák érzékenyek voltak a magas frekvenciákra, amilyeneket az utódaik produkálhattak. Weishampel szerint ez összhangban van a szülők és az utódok kommunikációjával.
A P. tubicen egy jól megőrzött, a P. walkeriénél bonyolultabb légutakkal rendelkező példányának számítógépes modellezése lehetővé tette annak a lehetséges hangnak a rekonstruálását, amelyet a gerince produkált. A főpálya 30 Hz körül rezonál, de a bonyolult szinusz anatómia csúcsokat és völgyeket okoz a hangban.
Hűtőfunkció
A címer nagy felülete és érhálózata hőszabályozó funkcióra is utal. P. E. Wheeler javasolta először 1978-ban ezt a felhasználást, mint az agy hűtésének módját. Teresa Maryańska és Osmólska nagyjából ugyanebben az időben szintén a hőszabályozást javasolta, Sullivan és Williamson pedig további érdeklődést mutatott. David Evans 2006-os értekezése a lambeosauruszok gerincfunkcióiról kedvezően fogadta az ötletet, legalábbis mint a gerinctágulás evolúciójának kezdeti tényezőjét.
A médiában
A Parasaurolophus egy nagyon népszerű dinoszaurusz. A Disney Dinoszaurusz című filmjében szerepelt a csorda tagjaként. A Dinoszauruszok összecsapása című híres dokumentumfilmben is többször szerepelt. A Prehistoric Park című népszerű tévéműsorban is feltűnt, ahol a nagy húsevők, a Deinosuchus és az Albertosaurus zsákmánya lett. A Jurassic Park filmekben is feltűnt néhányszor, az első filmben csordatagként, majd a másodikban vadászok fogságában, a harmadikban pedig a Corythosaurusszal együtt futva. és a Dweeb karaktere a Visszatértünk! A Dinosaur’s Story”, maga is egy Parasaurolophus.
A Parasaurolophus a Turokban is megjelenik, mint szelíd növényevő, aki általában nem ártalmas, de agresszív lehet, ha erősen provokálják.
A Power Rangersben is van egy Parasaurolophus zord: Dino Thunder és Power Rangers: Dino Charge.
A Parasaurolophus röviden megjelenik a Disney Pixar A jó dinoszaurusz című filmjének elején.
See Also
Parasaurolophus/Galéria
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, 229. oldal. Megjelent: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Chapter: Parasaurolophus, 678-684. oldal. Megjelent: 1997, McFarland & Co, in Jefferson, North Carolina. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull és Wright, Hadrosaurian Dinosaurs of North America, pp. 209-213.
- Brett-Surman, Michael K. és Wagner, Jonathan R. Carpenter, Kenneth (szerk.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, fejezet: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, pages 135-169. Megjelent, 2006, Indiana University Press, in Bloomington and Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; és Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; és Dodson, Peter (szerk.) The Dinosauria, 2nd edition, chapter: Hadrosauridae, 438-463. oldal. Megjelent: 2004, University of California Press, Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a crested trachodont dinosaurusok új neme és faja, 13. kötet, 1-32. oldal. Megjelent: 1922, University of Toronto Studies, Geology Series.
- 7,0 7,1 7,2 7,3 7,4 7,5 7,6 7,7 Sullivan, Robert M. és Williamson, Thomas E. A Parasaurolophus (Dinosauria: Hadrosauridae) új koponyája az új-mexikói Kirtland Formációból és a nemzetség felülvizsgálata, a sorozatból: New Mexico Museum of Natural History and Science Bulletin, 15., 1-52. oldal. Megjelent: 1999, New Mexico Museum of Natural History and Science, Albuquequeque, Új-Mexikó.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A hadrosaurian dinosaurian új faj az új-mexikói krétából, Journal of Paleontology, 35. kötet, 3. szám, 575-577. oldalon.
- Gilmore, Charles W., On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, 38. kötet, 43. szám, 29-48. oldal, Parks. Megjelent: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. és Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; és Dodson, Peter (szerk.) The Dinosauria, 1. kiadás, Chapter: Hadrosauridae, 534-561. oldal. Megjelent: 1990, University of California Press in Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., and Reisz, Robert R. 2007. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta, from the Journal of Vertebrate Paleontology, vol. 27 issue 2, on pages 373-393.
- Godefroit, Pascal, Shuqin Zan; and Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., a lambeosaurus dinosaurus az északkelet-kínai késő maastrichtiumból, a Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, pages 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Published by New Mexico Museum of Natural History and Science, in Albuquequeque, New Mexico. Oldalak 191-213.
- Liddell, Henry George és Robert Scott, 1980. A Greek-English Lexicon (rövidített kiadás) Kiad: Oxford University Press az Egyesült Királyságban. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. és Evans, David C., Currie, Phillip J. és Koppelhus, Eva (szerk.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, Chapter: Ornithischian Dinosaurs. 2005, Kiadva: Indiana University Press, Bloomingtonban. Oldalak 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; and Dupuis, Kevin, 2007. Egy fiatalkorú Parasaurolophus agykoponya az albertai Dinosaur Provincial Parkból, megjegyzésekkel a nemzetség gerincének ontogeneziséről. Journal of Vertebrate Paleontology, vol. 27, issue 3 pages 642-650.
- Currie, Phillip J; and Russell, Dale A. “Geographic and stratigraphic distribution of dinosaur remains” in Dinosaur Provincial Park, p. 553. o.
- Weishampel, David B. (1990). “Dinosaur Distribution”, in The Dinosauria (1st), pp. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). “Dinosaur Distribution”, in The Dinosauria (2.), pp. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, from the Nova Acta Regia Societas Scientarum Upsaliensis, series 4, vol. 7, issue 5. (németül). Oldalak 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, kiadó: Cassell Ltd. 1979, 5. kiadás, Londonban. ISBN 0-304-52257-0 883. oldal.
- Weishampel, David B. és Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) Utahból, a Journal of Paleontology 53. évfolyamának 6. számából, 1422-1427. oldal.
- Eberth, David A. 2005. “A geológia”, in: Dinosaur Provincial Park, 54-82. oldal.
- Braman, Dennis R. és Koppelhus, Eva B. 2005. “Campanian palynomorphs”, in Dinosaur Provincial Park, pp. 101-130.
- Russell, Dale A. An Odyssey in Time: Dinosaurs of North America, 1989. Kiadó: NorthWord Press, Minocqua, Wisconsin. ISBN 1-55971-038-1. 160-164. oldal.
- Bakker, Robert T. 1986. A dinoszauruszok eretnekségei: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, Kiadó: William Morrow, New York. ISBN 0-8217-2859-8 194. oldal.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: Eredeti és lenyűgöző betekintés a dinoszauruszok birodalmának életébe, fejezet: Hadrosauridák II. 1985. Kiadja a Crescent Books, New Yorkban. 122-127. oldal. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Orrüreg homológiák és a koponyacsúcs funkciója a lambeosaurus dinoszauruszoknál, a Journal of Paleobiology, vol. 32, issue 1, Pages 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurian , from the Journal of Paleobiology, volume one, issue one, Pages 21-43.
- Romer. Alfred Sherwood, 1933. Gerinces paleontológia, a University of Chicago Press kiadványából, a 491. oldalon.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, az Orion folyóirat (München) 2. kötetéből, 525-532. oldal (német nyelven).
- Sternberg, Charles M. 1935. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous from the Journal of the Canada Department of Mines Bulletin (Geological Series), volume 77, issue 52, on pages 1-37.
- Colbert, Edwin H. The Dinosaur Book: Az uralkodó hüllők és rokonaik, kiadta 1945-ben az American Museum of Natural History, Man and Nature Publications, 14, New York. 156. oldal.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas a Jarbuch Naturwissenschaften folyóirat 12. kötetének 36. számában a 709-716. oldalon. (németül) 1924.
- Osmólska, Halszka, 1979. Orrmelléki sómirigyek a dinoszauruszoknál, az Acta Palaeontologica Polonica folyóirat 24. évfolyamának 205-215. oldaláról.
- 36.0 36.1 Ostrom, John H., 1962. The cranial crests of hadrosaurian dinosaurian, from the Journal of Postilla, vol. 62, pages 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, kiadó: Master Books, in Green Forest. ISBN 0-89051-165-9 82. oldal.
- 38.0 38.1 Weishampel, David B., 1981. A lambeosaurus dinoszauruszok (Reptilia:Ornithischia) potenciális vokalizációjának akusztikai elemzése, a Journal of Paleobiology, vol. 7, issue 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, from the Journal of Paleontology, vol. 55, issue 5, pp. 1046-1057.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur, a Sandia National Laboratories-tól (1997-12-05). Retrieved on January 20th, 2009.
- Diegert, Carl F. és Williamson, Thomas E., 1998. A lambeosaurine hadroszaurusz Parasaurolophus tubicen digitális akusztikai modellje a Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. és Williamson, Thomas E., 1996. A Parasaurolophus (long-crested form) új koponyája Új-Mexikóból: külső és belső (CT felvételek) jellemzők és funkcionális vonatkozásaik, a Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborate CNS cooling structure in large dinosaurs Journal of Nature, vol. 275, on pp. 441-443.
- Maryańska, Teresa és Osmólska, Halszka, 1979-ben. Aspects of hadrosaurian cranial anatomy, a Journal of Lethaia, vol. 12, on pp. 265-273.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur; from Sandia National Laboratories.
- Restorations of P. walkeri and P. cyrtocristatus, to the same scale, by Scott Hartman; at Skeletal Drawing.com.
- Parasaurolophus page on Enchanted Learning.com
.