

パラサウロフス(発音:/ˌpærəsɔˈrɒləfəs/ PARR-)ə-saw-ROL-ə-fəsです。 一般に、/ˌpærəˌsɔrˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs とも呼ばれる。 Saurolophusを指して「トカゲに近い」という意味)は、現在の北米の白亜紀後期、約7600万年前に生息していた鳥脚類恐竜の一属です。 二足歩行と四足歩行の両方をする草食動物であった。 3種が確認されている。 P. walkeri(原種)、P. tubicen、そして短頭種のP. cyrtocristatusの3種が確認されている。 カナダのアルバータ州、アメリカのニューメキシコ州、ユタ州から遺存することが確認されている。
パラサウロロフスはハドロサウルス類で、白亜紀の恐竜の中でも頭部の装飾が多様なことで知られている科に属しています。 この属は、大きく精巧な頭蓋紋を持つことで知られており、最も大きいものでは、頭蓋骨から上方および後方に突き出た長い湾曲した筒を形成しています。 最も近縁と思われる中国のチャロノサウルスも同様の頭蓋骨を持ち、同様の紋章を持っていた可能性がある。 この紋章については科学者たちの間で多くの議論が交わされてきたが、主な機能としては、種や雌雄の視覚的認識、音響共鳴、体温調節などが挙げられるというのが一致した見解である。
パラサウロロフスは最もよく知られている恐竜のひとつで、恐竜関連のテレビ番組、映画、本、グッズなどのメディアで頻繁に描かれている。 アリス・ペリー
Description

Parasaurolophus walkeri の大きさを人間で比べると
多くの恐竜と同様に Parasaurolophus も骨格は不完全にしかわかっていません。 ウォーケリのタイプ標本の全長は9.5メートルと推定されています。 一方、P. tubicenの頭骨は2.0メートル以上あり、より大型の動物であることがわかる。 体重は2.5トン(2.7トン)と推定されている。 知られている単一の前肢はハドロサウルス類としては比較的短く、短いが幅の広い肩甲骨を持つ。 P. walkeriの大腿骨の長さは103センチメートルで、他のハドロサウルス類と比較すると、その長さの割には頑丈である。 上腕骨や骨盤も重く作られている。
他のハドロサウルス類と同様に、2本足でも4本足でも歩くことができた。 おそらく4本足で食物を採ることを好んだが、2本足で走ったのであろう。 脊椎骨の神経棘はランベオサウルス類によく見られるように高く、腰の上で最も高くなり、背の高さを増しています。
Parasaurolophus walkeri with scalation detail.
最も顕著な特徴は頭頂で、これは後頭部に突き出ていて前顎と鼻骨から構成されている。 P. walkeriのタイプ標本では、クレストが背中に当たる付近の神経棘に切り欠きがありますが、これはこの個体特有の病理である可能性があります。 属名を命名したウィリアム・パークスは、頭頂部から切り欠き部まで靭帯が走り、頭部を支えているのではという仮説を立てた。 その可能性は低いと思われますが、パラサウロロフスの復元には、頂部から首にかけての皮弁が用いられることがあります
頂部は中空で、それぞれの鼻孔から頂部の端に至るまで、方向を反転して頂部から頭蓋骨に戻る明確な管を持っていました。 管はP. walkeriでは最も単純で、P. tubicenではより複雑で、ある管は盲目で、他の管は合流して分離していた。
分類
パラサウロロフスはその名の通り、表面的に紋が似ているため、当初はサウロロフスに近縁と考えられていました。 しかし、すぐにハドロサウルス類のランベオサウルス亜科の一員-サウロロフスはハドロサウルス類-として再評価された。 通常、ヘルメット紋を持つコリトサウルス、ハイパクロサウルス、ランベオサウルスとは別系統のランベオサウルス類と解釈される。 最も近縁なのは、中国北東部のアムール地方に生息するチャロノサウルスで、同様の頭骨をもつランベオサウルス類(ただし、まだ完全な紋はない)であり、両者はパラサウロロフィニ類のクレードを形成する可能性がある。 P. cyrtocristatusは、短く丸い紋を持ち、既知の3種のParasaurolophusの中で最も基底にある種か、P. tubicenのサブアダルトまたはメス標本であるかもしれない。
歴史
発見と命名
ワルシャワにあるParasaurolophus walkeriの不完全なタイプ標本の鋳型
「トカゲに近い」という意味のParasaurolophusという名前は、ギリシャ語のpara/παρα「そば」「近く」、saurus/σαυρος「トカゲ」と lophos/λοφος 「紋」から由来しています。 これは、1920年にトロント大学の野外調査団がカナダ・アルバータ州のレッドディア川沿いのサンドクリーク付近で発見した、尾の大部分と膝下の後肢を失った頭蓋骨と部分骨格であるROM 768を基にしています。 これらの岩石は現在、カンパニア紀白亜紀上期のダイナソーパーク層として知られている。 ウィリアム・パークスは、この標本をロイヤル・オンタリオ博物館の評議員会会長であるバイロン・エドモンド・ウォーカー卿にちなんで、P. walkeriと命名した。 パラサウロロフスの遺体はアルバータ州では珍しく、他に(おそらく)恐竜公園層から部分的な頭骨が出土したのみで、恐竜公園標本の3体は頭骨を欠き、同属である可能性がある。 いくつかの動物リストには、マーストリヒト後期のモンタナ州ヘル・クリーク層でP. walkeriの可能性があるとの記載があるが、この岩石層はマーストリヒト後期の岩石層である。 1921年にCharles H. Sternbergがニューメキシコ州San Juan郡のやや若いKirtland層から頭骨の一部(PMU.R1250)を採集した。 この標本はスウェーデンのウプサラに送られ、Carl Wimanによって1931年に第2種 P. tubicenとして記載された。 この種はラテン語のtǔbǐcěn “trumpeter “を語源としている。 1995年にニューメキシコ州で2頭目のほぼ完全なP. tubicenの頭骨(NMMNH P-25100)が発見された。 この頭骨のCT(コンピュータ断層検査)を用いて、Robert SullivanとThomas Williamsonは1999年にこの属を単行本化し、解剖学、分類学、紋章の機能などの側面をカバーしました。
John Ostromは1961年にニューメキシコ州のもう一つの良い標本 (FMNH P27393) をP. cyrtocristatusとして報告した。 この標本には、短く丸い紋章を持つ頭蓋骨の一部と、足、首、尾の一部を除く頭蓋後部の骨格の大部分が含まれている。 種名はラテン語のcurtus「短くなった」とcristatus「紋章のある」に由来する。 この標本はフルーツランド層の上部か、その上にあるカートランド層の基部で発見された。 1979年にDavid B. WeishampelとJames A. Jensenがユタ州ガーフィールド郡のカンパニアン時代のカイパロウィッツ層から同様の紋章をもつ部分頭骨(BYU 2467)を報告し、本種の生息域が拡大されることになった。 それ以来、ユタ州ではP. cyrtocristatusの短い/丸い紋章の形態を持つ別の頭蓋骨が見つかっている。
種
Parasaurolophus walkeri 頭蓋骨キャスト、ロンドン自然史博物館、P.
Parasaurolophus cyrtocristatus 頭蓋骨キャスト、フィールド自然史博物館
タイプ種P. walkeriは、アルバータから1つの標本をもって知られています。 また、P. cyrtocristatusとは、紋が長く、丸みがなく、上腕が前腕より長い点で異なる。
ニューメキシコ州のP. tubicenは、少なくとも3個体の遺体から知られている。
ニューメキシコ州とユタ州のP. cyrtocristatusは、3つの可能性のある標本から知られている。 ニューメキシコ州とユタ州のP. cyrtocristatusは3つの可能性のある標本から知られており,最も小型の種で,紋は短く丸みを帯びる。 その小ささと紋の形から、ほぼ同時期にニューメキシコ州の同じ地層から産出されたP. tubicenの幼体あるいは雌であるとする研究者がいる。 トーマス・ウィリアムソンによると、P. cyrtocristatusの型板はP. tubicenの約72%の大きさで、他のランベオサウルス類が紋章に決定的な性的二型を示し始めるとされるサイズ(大人のサイズの約70%)とほぼ同じであると言うことである。
Paleobiology
Paleoecology
Parasaurolophus cyrtocristatusを攻撃するゴルゴサウルス(左)。
Parasaurolophus walkeriは恐竜公園層から出土し、角のあるセントロサウルス、スティラコサウルス、カスモサウルスなどのよく知られた恐竜を含む先史時代の動物の多様でよく知られた動物群の一員であった。 ティラノサウルス類のゴルゴサウルス、鎧竜のエドモントニア、ユオプロケファルスなどである。 この動物群の中では珍しい構成員であった。 恐竜公園層は、西部内陸海道の西方への侵犯に伴い、時間の経過とともに海洋条件の影響を受け、より湿潤になった河川や氾濫原の低レリーフ設定と解釈される。 気候は現在のアルバータ州より暖かく、霜はなかったが、雨季と乾季があった。
ニューメキシコの種は、大型竜脚類のアラモサウルス、アヒル嘴のクリトサウルス、角のあるペンタケラトプス、装甲類のノドケファロサウルス、サウルニソレス、そして現在名前がないティラノサウルス類と環境を共有していたようである。 カートランド層は、西内部海路の後退後に出現した河川氾濫原と解釈されている。
摂食
ハドロサウルス類のパラサウロロフスは、大型の二足歩行/四足歩行草食動物で、咀嚼に似た研削運動を可能にする高度な頭蓋骨で植物を食べていました。 歯は常に交換され、数百本の歯が入った歯列に詰め込まれていたが、常に使用されていたのはそのうちのほんの一握りであった。 嘴を使って植物を収穫し、それを頬のような器官で顎に保持した。 餌は地上から4メートル上空までの高さにあっただろう。 ボブ・バッカーが指摘したように、ランベオサウルス類はハドロサウルス類よりもくちばしが細いので、パラサウロロフスとその近縁種はくちばしが広く、紋章のない同種の動物よりも選択的に餌を食べることができたと思われる。 現在では、種や雌雄を識別するための視覚的表示、コミュニケーションのための音の増幅、体温調節などの機能があったのではないかと考えられている。 8107>
種や成長段階の違い
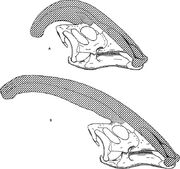
Parasaurolophus cyrtocristatus (a) と Parasaurolophus walkeri (b) の鼻紋を比較した図です。
他のランベオサウルス類と同様に、パラサウロロフスの頭蓋紋は年齢とともに変化し、成体では性差のある特徴であったと考えられています。 このような区別の観点からランベオサウルス類の紋章を記述した最初の研究者の一人であるJames Hopsonは、小さな紋章を持つP. cyrtocristatusをP. tubicenの雌型であると示唆した。 トーマス・ウィリアムソン(Thomas Williamson)は幼体であるとした。 どちらの仮説も広く受け入れられることはなかった。 良質の頭骨が6個、幼体の脳槽が1個しか知られていないので、さらなる資料があれば、これらの関係の可能性を明らかにすることができるだろう。 ウィリアムソン氏は、いずれにせよ、パラサウロロフスの幼体はおそらくP. cyrtocristatusのように小さく丸い紋を持っていて、成熟に近づくにつれて早く成長すると指摘している。 以前はランベオサウルスとされ、現在はパラサウロロフスのものとされた幼獣の脳容器の最近の再研究により、幼獣に小さな管状紋が存在した証拠が得られています。 この標本には、成体標本に見られるものと似ているがより小さい、前頭骨の上方への小さな膨らみが残っている。成体では、前頭骨は紋章の基部を支える土台を形成していたのである。 この標本は、パラサウロロフスの紋章の成長と幼体個体の顔の輪郭が、コリトサウルス・ハイパクロサウルス・ランベオサウルスモデルと異なることを示す。これはパラサウロロフスの紋章には、他の3種のランベオサウルス類の紋章上部を構成する薄い骨の「コックスコム」がないことが一因であると考えられる。
機能に関する仮説の否定
初期の多くの提案は、1960年代までよく見られたハドロサウルス類が水陸両用であるという仮説に従って、水中生活への適応に焦点を当てたものであった。 アルフレッド・シャーウッド・ロマーはシュノーケルの役割を、マーティン・ウィルファースは呼吸器や食物採集のための移動口吻のアタッチメントを、チャールズ・M・スタンバーグは肺に水が入らないようエアトラップの役割を、ネッド・コルバートは水中に長時間いるための空気貯留の役割を果たしたと提案しています
他の提案はより物理的な性質を持っています。 前述のように、ウィリアム・パークスは、靭帯や筋肉で脊椎骨と結合し、頭を動かしたり支えたりするのに役立っていると提案した。 また、オテニオ・アベルは同種族間の戦闘で武器として使用することを提案し、アンドリュー・ミルナーはヒクイドリの兜の紋章(「カスク」と呼ばれる)のように葉をよけるものとして使用することを提案した。 さらに、特殊な器官を収容することを主要な機能とする案もあった。 ハルシュカ・オスモルスカは塩類腺を収容していると示唆し、ジョン・オストロムは嗅覚組織のための領域を拡大し、明らかな防御能力を持たないランベオサウルス類の嗅覚をはるかに向上させたと示唆した。 8107>
これらの仮説のほとんどは、信用されなかったり、否定されたりしています。 たとえば、紋章の先端にはシュノーケリングの機能を果たす穴がない。 口吻のための筋肉の傷もなく、くちばしのある動物が口吻を必要とするかどうかも疑わしい。 提案されたエアロックとして、水を防ぐことはできなかっただろう。 パラサウロロフスのような大きさの動物にとって、提案された空気溜めは不十分であったろう。 他のハドロサウルス類は大きな頭部を持ち、靭帯を支えるための大きな中空紋を必要としなかった。 また、どの提案も、紋章がなぜそのような形をしているのか、なぜ他のランベオサウルス類は見た目は大きく異なるが同様の機能を果たす紋章を持っているのか、紋章のないハドロサウルス類や強固な紋章を持つハドロサウルス類はどのようにしてそのような機能なしでやっていけるのか、なぜ一部のハドロサウルス類は強固な紋章を持っていたのかを説明しないままになっている。 これらの考察は、塩類腺仮説や嗅覚仮説など、動物にすでに存在するシステムの能力を高めることに基づく仮説に特に影響を与え、これらが紋章の主要機能ではなかったことを示すものである。 さらに、ランベオサウルス類の鼻腔の研究から、嗅覚神経とそれに対応する感覚組織は、顎堤の鼻腔の部分の外側に多く存在することがわかり、顎堤の拡張は嗅覚とはほとんど関係ないことがわかった。walkeri3.jpg Life restoration of Parasaurolophus walkeri with hypothetical skin frill and stripes on its crest.
instead, social and physiological functions are more supported as the function(s) with visual and auditory identification and communicationに焦点を当てて、紋章の機能について検討するようになった。 紋章は大型の物体であるため、視覚信号としての価値が明確であり、同時代の動物とは一線を画している。 ハドロサウルス類の眼窩が大きく、眼球に硬化輪があることから、視力が鋭く、昼行性であり、この動物にとって視力が重要であったことを示す証拠である。 もし、一般的に描かれているように、紋章から首や背中まで皮膚のフリルが伸びていたら、提案された視覚ディスプレイはもっと派手なものになったはずである。 他のランベオサウルス類の頭蓋骨で示唆されているように、パラサウロロフスの紋章は、種の識別(コリトサウルスやランベオサウルスと分けるなど)と、形や大きさから雄と雌を判断することの両方を可能にしたと思われます。jpg P. tubicenの紋章の鼻腔のコンピュータモデル。
しかしながら、紋章の外見と鼻腔の複雑な内部解剖は一致せず、内部空間の使用は別の機能で説明できると考えられる。 カール・ワイマンは1931年に、鼻腔がクラムホーンのような聴覚的な信号機能を果たすと最初に提案しました。ホプソンとデヴィッド・B・ワイシャンペルは、1970年代と1980年代にこの考えを再検討しました。 ホプソンは、ハドロサウルス類が強い聴覚を持っていたことを示す解剖学的な証拠があることを発見した。 近縁種のコリトサウルスには、細長いアブミ骨(爬虫類の耳の骨)がある例が少なくとも1つあり、鼓膜のスペースが広いことと合わせて、中耳が敏感だったことを示唆しているのである。 さらに、ハドロサウルス類のラゲーナはワニのように細長く、内耳の聴覚部分がよく発達していたことがわかる。 Weishampelは、P. walkeriは48から240Hz、P. cyrtocristatus(紋章型の幼生と解釈)は75から375Hzの周波数を出すことができると示唆した。 また、ハドロサウルス類の内耳がワニの内耳と似ていることから、彼はハドロサウルス類の成体は、彼らの子孫が出すような高い周波数に敏感であるとも提唱している。 Weishampel によると、これは親と子がコミュニケーションをとることと一致します。
P. walkeri よりも複雑な空気通路を持つ P. tubicen の保存状態のよい標本のコンピューターモデル化により、その頂部が出すと考えられる音の復元が可能になりました。
冷却機能
頂部の表面積が大きく、血管が多いことも体温調節機能を示唆している。 P.E. Wheelerは1978年に、脳を冷やす方法としてこの用途を初めて提案しました。 Teresa MaryańskaとOsmólskaもほぼ同時期に体温調節機能を提案し、SullivanとWilliamsonがさらに関心を持った。 David Evansの2006年のlambeosaurine crest functionの議論は、少なくともcrest expansionの進化の初期要因として、この考えに好意的だった。
In The Media
Parasaurolophusは非常に人気のある恐竜である。 映画「ディズニーダイナソー」にも群れの一員として出てきました。 また、有名なドキュメンタリー映画「恐竜大激突」にも何度か登場しました。 人気テレビ番組「プレヒストリック・パーク」にも登場し、大型肉食獣のデイノサクスやアルベルトサウルスの餌食となった。 映画『ジュラシック・パーク』にも登場し、第1作では群れの一員として、第2作ではハンターに捕らえられ、第3作ではコリトサウルスと一緒に走っている姿が描かれている。 8107>
パワーレンジャーにもパラサウロロフスのゾードが登場します。 ディノサンダーとパワーレンジャー。 8107>
Parasaurolophus はディズニー・ピクサー映画「The Good Dinosaur」の冒頭に少しだけ登場します。
See Also
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, page 229. 発行:1942年、アメリカ地質学会、Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Chapter.3: Parasaurolophus, pages 678-684. 1997, McFarland & Co, in Jefferson, North Carolina.発行。 ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull and Wright, Hadrosaurian Dinosaurs of North America, pp.209-213.による。
- Brett-Surman, Michael K. and Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, chapter: カンパニアンとマーストリヒト北アメリカのハドロサウルス類の肩甲骨の解剖学、ページ135-169。 発行, 2006, インディアナ大学出版会, in Bloomington and Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; And Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) The Dinosauria, 2nd edition, chapter: ハドロサウルス科、438-463ページ。 2004年、カリフォルニア大学出版局(バークレー)発行。 ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur.第13巻、1-32ページ. 発行:1922年、トロント大学研究、地質学シリーズ。
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. and Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, from the series New Mexico Museum of Natural History and Science Bulletin, 15, pages 1-52. 発行:1999年、ニューメキシコ州アルバケケのニューメキシコ自然史・科学博物館。
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, Journal of Paleontology, Volume 35, 3rd issue, on pages 575-577.
- Gilmore, Charles W., On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, volume 38, issue 43, pages 29-48, パークス.「ステファノサウルス属について」『古生物学研究』第38巻第4号. 公開:1924年、カナダ鉱山省地質調査所紀要(地質学シリーズ)
- Weishampel, David B. and Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) The Dinosauria, 1st edition, Chapter.The Dinosauria, Chapter.The David B. and Horszka, Dodson, Peter, December 1924, Canada Department of Mines, Geological Survey Bulletin (Geological Series), Chapter.1, 1924: ハドロサウルス科、534-561ページ。 1990年、カリフォルニア大学出版局(バークレー)発行。 ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., and Reisz, Robert R. 2007.日本学術振興会特別研究員(DC)。 この論文では、”Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta”, Journal of Vertebrate Paleontology, vol.27 issue 2, on pages 373-393.に掲載されたLambeosaurusの解剖と関係性について述べています。
- Godefroit, Pascal, Shuqin Zan; and Liyong Jin. 2000. また,”Science de la Terre et des Planètes” Vol.330 の 875-882 ページに,中国東北部の Maastrichtian 後期のランベオサウルスである Charonosaurus jiayinensis n. g., n. sp.の記載がある.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. ニューメキシコ州サンフアン盆地のハドロサウルス科(恐竜目鳥綱)についてのレビュー Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 ニューメキシコ州アルバケキューのニューメキシコ自然史科学博物館が出版したもので、ニューメキシコの自然史科学博物館は、その中に、ハドロサウルス科(DINOSURIA, ORGNISCHIA)を含む。 191-213ページ。
- Liddell, Henry George and Robert Scott, 1980. A Greek-English Lexicon (Abridged Edition) 発行。 オックスフォード大学出版局(英国)。 ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. and Evans, David C., Currie, Phillip J., and Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed(壮大な古代の生態系が明らかになった), Chapter: 鳥盤類恐竜. 2005年、発行。 インディアナ大学出版、ブルーミントン。 また、このような研究成果をもとに、「恐竜の生態系を解明する」と題したシンポジウムを開催しました。 また、このような俯瞰的な視点に立つことで、より高度な分析が可能となる。 また、このような場合にも、「震災の影響」を考慮する必要があります。
- Currie, Phillip J; and Russell, Dale A. “Geographic and stratigraphic distribution of dinosaur remains” in Dinosaur Provincial Park, p. 553.
- Weishampel, David B. (1990). “Dinosaur Distribution”, The Dinosauria (1st), pp.63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004).日本産恐竜の分布(第1回). “Dinosaur Distribution”, in The Dinosauria (2nd), pp.517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, from the Nova Acta Regia Societas Scientarum Upsaliensis, series 4, vol.7, issue 5. (ドイツ語)。 1-11ページ。
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, published by Cassell Ltd. (英語). 1979年、第5版、ロンドンにて。 ISBN 0-304-52257-0 883ページ。
- Weishampel, David B. and Jensen, James A. 1979. ユタ州産Parasaurolophus (Reptilia: Hadrosauridae), Journal of Paleontology, vol.53, issue 6, pages 1422-1427より。
- Eberth, David A. 2005. “The geology”, in Dinosaur Provincial Park, pp.54-82.
- Braman, Dennis R., and Koppelhus, Eva B. 2005. 「カンパニアン植物形態学」(恐竜州立公園,pp.101-130.所収).
- Russell, Dale A. An Odyssey in Time: 北アメリカの恐竜たち, 1989. 出版社。 ウィスコンシン州ミノクア所在の NorthWord Press, 。 ISBN 1-55971-038-1 160-164ページ。
- Bakker, Robert T. 1986. 恐竜の異端児。 恐竜とその絶滅の謎を解く新説、ウィリアム・モロー社、ニューヨークで出版。 ISBN 0-8217-2859-8 194ページ。
- 27.0 27.1 27.2 ノーマン・デビッド・B. The Illustrated Encyclopedia of Dinosaurs(恐竜図鑑)。 An Original and Compelling Insight into Life in the Dinosaur Kingdom(恐竜王国における生命への独創的で説得力のある洞察), chapter: ハドロサウルス類II. 1985. 1985年、ニューヨークのクレセント・ブックス発行。 ページ122-127。 ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. ランベオサウルス類における鼻腔の相同性と頭蓋堤の機能」『古生物学雑誌』32巻1号、109-125頁より。
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. ハドロサウルス類恐竜の頭蓋表示構造の進化 、古生物学雑誌、第1巻、第1号、21-43ページより。
- ロマー. アルフレッド・シャーウッド、1933年 Vertebrate Paleontology, from the University of Chicago Press, on page 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, from the Journal of Orion (Munich), vol. 2. pp.525-532(ドイツ語).
- Sternberg, Charles M. 1935. 白亜紀後期のベリーリバー・シリーズのフード付きハドロサウルス カナダ鉱山局紀要(地質学シリーズ)第77巻52号1〜37頁より。
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, 1945年アメリカ自然史博物館発行、Man and Nature Publications, 14, in New York.より。 156ページ
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas Journal of Jarbuch Naturwissenschaften, volume 12, issue 36, on pages 709-716. (独) 1924.
- Osmólska, Halszka, 1979. 恐竜の鼻塩類腺、Acta Palaeontologica Polonica誌24巻、205-215頁より。
- 36.0 36.1 Ostrom, John H., 1962. ハドロサウルス類恐竜の頭蓋紋(『ポスティラ誌』第62巻、1-29頁より).
- Gish, Duane T.、1992. Dinosaurs by Design, マスターブックス刊、グリーンフォレストにて。 ISBN 0-89051-165-9 82ページ。
- 38.0 38.1 Weishampel, David B., 1981. ランベオサウルス類(爬虫類:鳥綱類)の潜在的発声の音響学的解析」『古生物学雑誌』第7巻第2号、252-261頁より。
- Weishampel David B., 1981. ハドロサウルス類の鼻腔:比較解剖学とホモロジー」『古生物学雑誌』55巻5号、1046-1057頁。
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur, by the Sandia National Laboratories (1997-12-05). 2009 年 1 月 20 日に取得。
- Diegert, Carl F. and Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen from the Journal of Vertebrate Paleontology, vol.18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. and Williamson, Thomas E., 1996. ニューメキシコ州産パラサウロロフスの新頭骨:外形・内部(CTスキャン)の特徴とその機能的意味」『脊椎動物古生物学雑誌』16巻3号付録、pp.68Aより。
- Wheeler, P.E., 1978. 大型恐竜における精巧なCNS冷却構造 Journal of Nature, vol.275, on pp.441-443.
- Maryańska, Teresa and Osmólska, Halszka, in 1979. Aspects of hadrosaurian cranial anatomy, from the Journal of Lethaia, vol.12, on pp.265-273.
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur; from Sandia National Laboratories.
- Restoreations of P. walkeri and P. cyrtocristatus, by Scott Hartman; at Skeletal Drawing.com.
- Enchanted Learning.com
Parasaurolophus page on P. cortocristatus from Sandia Nation Laboratories.Online.com