

Parasaurolophus (uitgesproken als /ˌpærəsɔˈrɒləfəs/ PARR-ə-saw-ROL-ə-fəs, gewoonlijk ook /ˌpærəˌsɔrəˈloʊfəs/ PARR-ə-SAWR-ə-LOH-fəs; wat “bijna-kuifhagedis” betekent in verwijzing naar Saurolophus) is een geslacht van ornithopode dinosauriërs uit de Late Krijtperiode van wat nu Noord-Amerika is, ongeveer 76-73 miljoen jaar geleden. Het was een herbivoor die zowel als twee- en viervoeter liep. Er zijn drie soorten erkend: P. walkeri (de typesoort), P. tubicen, en de kortgekuifde P. cyrtocristatus. Er zijn overblijfselen bekend uit Alberta, Canada, en New Mexico en Utah, USA. Hij werd voor het eerst beschreven in 1922 door William Parks aan de hand van een schedel en een gedeeltelijk skelet in Alberta.
Parasaurolophus is een hadrosauriër, die deel uitmaakt van een diverse familie van dinosauriërs uit het Krijt die bekend staan om hun reeks bizarre hoofdversieringen. Dit geslacht is bekend om zijn grote, uitgebreide schedelkam, die op zijn grootst een lange gebogen buis vormt die omhoog en naar achteren uit de schedel steekt. Charonosaurus uit China, die misschien zijn naaste verwant was, had een gelijkaardige schedel en mogelijk een gelijkaardige kuif. Over de kuif is door wetenschappers veel gediscussieerd; de consensus is dat de belangrijkste functies visuele herkenning van zowel soort als geslacht, akoestische resonantie, en thermoregulatie omvatten. Het is een van de zeldzaamste eendenbekken, bekend van slechts een handvol goede exemplaren.
Parasaurolophus is een van de meest herkende dinosaurussen, en wordt vaak afgebeeld in dinosaurus-gerelateerde televisieprogramma’s, films, boeken, merchandise en andere media. Alice Perry
Beschrijving

De grootte van Parasaurolophus walkeri vergeleken met een mens.
Zoals bij de meeste dinosauriërs het geval is, is het skelet van Parasaurolophus onvolledig bekend. De lengte van het type-exemplaar van P. walkeri wordt geschat op 9,5 meter (31 ft). Zijn schedel is ongeveer 1,6 meter lang, inclusief de kuif, terwijl de typeschedel van P. tubicen meer dan 2,0 meter lang is, wat op een groter dier wijst. Zijn gewicht wordt geschat op 2,5 ton (2,7 ton). Zijn enige bekende voorpoot was relatief kort voor een hadrosauride, met een kort maar breed schouderblad. Het dijbeen meet 103 centimeter bij P. walkeri en is robuust voor zijn lengte in vergelijking met andere hadrosauriden. De botten van de bovenarm en het bekken waren ook zwaar gebouwd.
Zoals andere hadrosauriden kon hij zowel op twee als op vier poten lopen. Hij gaf er waarschijnlijk de voorkeur aan op vier poten voedsel te zoeken, maar liep op twee poten. De neurale stekels van de wervels waren hoog, zoals gebruikelijk was bij lambeosaurines; het hoogst boven de heupen, verhoogden zij de hoogte van de rug. Van P. walkeri zijn huidindrukken bekend, die uniforme knobbelachtige schubben vertonen, maar geen grotere structuren.
Parasaurolophus walkeri met scalatie-detail.
Het meest opvallende kenmerk was de craniale kam, die aan de achterzijde van de kop uitstak en bestond uit de premaxilla en neusbeenderen. Het type-exemplaar van P. walkeri heeft een inkeping in de neurale stekels nabij de plaats waar de kam de rug zou raken, maar dit kan een pathologie zijn die eigen is aan dit individu. William Parks, die het genus zijn naam gaf, veronderstelde dat een ligament van de kam naar de inkeping liep om de kop te ondersteunen. Hoewel dit idee onwaarschijnlijk lijkt, wordt Parasaurolophus soms gerestaureerd met een huidflap van de kam naar de nek.
De kam was hol, met duidelijke buizen die van elk neusgat naar het einde van de kam leidden, alvorens in omgekeerde richting terug te gaan langs de kam en in de schedel. De buizen waren het eenvoudigst bij P. walkeri, en complexer bij P. tubicen, waar sommige buizen blind waren en andere elkaar ontmoetten en scheidden. Terwijl P. walkeri en P. tubicen lange kuiven hadden met slechts een lichte kromming, had P. cyrtocristatus een korte kuif met een meer cirkelvormig profiel.
Classificatie
Zoals de naam al aangeeft, werd Parasaurolophus aanvankelijk geacht nauw verwant te zijn aan Saurolophus vanwege zijn oppervlakkig gelijkende kuif. Hij werd echter al snel geherwaardeerd als lid van de lambeosaurine onderfamilie van de hadrosauriden – Saurolophus is een hadrosaurine. Hij wordt gewoonlijk beschouwd als een afzonderlijke tak van de lambeosauriërs, te onderscheiden van de met helmkuif gekuifde Corythosaurus, Hypacrosaurus en Lambeosaurus. Zijn nauwst bekende verwant schijnt Charonosaurus te zijn, een lambeosaurine met een soortgelijke schedel (maar nog geen volledige kuif) uit het Amoer-gebied van Noordoost-China, en de twee vormen wellicht een clade Parasaurolophini. P. cyrtocristatus, met zijn korte, rondere kam, kan de meest basale van de drie bekende Parasaurolophus-soorten zijn, of het kan gaan om subadulte of vrouwelijke specimens van P. tubicen.
Geschiedenis
Ontdekking en naamgeving
Geboorte van het incomplete Parasaurolophus walkeri type-exemplaar in Warszawa.
De naam Parasaurolophus betekent “nabij kuifhagedis” en is afgeleid van het Griekse para/παρα “naast” of “nabij”, saurus/σαυρος “hagedis” en lophos/λοφος “kuif”. Het is gebaseerd op ROM 768, een schedel en gedeeltelijk skelet waarbij het grootste deel van de staart en de achterpoten onder de knieën ontbraken, die in 1920 door een veldploeg van de Universiteit van Toronto werd gevonden in de buurt van Sand Creek langs de Red Deer River in Alberta, Canada. Deze gesteenten staan nu bekend als de Campanien-tijdperk Boven-Krijt Dinosaur Park Formatie. William Parks noemde het specimen P. walkeri ter ere van Sir Byron Edmund Walker, voorzitter van de Board of Trustees van het Royal Ontario Museum. Parasaurolophus-resten zijn zeldzaam in Alberta, met slechts één andere gedeeltelijke schedel uit (waarschijnlijk) de Dinosaur Park Formation, en drie Dinosaur Park-specimens zonder schedel, die mogelijk tot het genus behoren. In sommige faunalijsten wordt melding gemaakt van mogelijk P. walkeri-materiaal in de Hell Creek Formation van Montana, een gesteente-eenheid van laat-Maastrichtien ouderdom. Dit voorkomen wordt niet vermeld door Sullivan en Williamson in hun bespreking van het genus in 1999, en is elders niet verder uitgewerkt.
In 1921 vond Charles H. Sternberg een gedeeltelijke schedel (PMU.R1250) uit wat nu bekend staat als de iets jongere Kirtland Formatie in San Juan County, New Mexico. Dit exemplaar werd opgestuurd naar Uppsala, Zweden, waar Carl Wiman het in 1931 beschreef als een tweede soort, P. tubicen. Het specifieke epitheton is afgeleid van het Latijnse tǔbǐcěn “trompetter”. Een tweede, bijna complete schedel van P. tubicen (NMMNH P-25100) werd in 1995 in New Mexico gevonden. Met behulp van computertomografie van deze schedel gaven Robert Sullivan en Thomas Williamson het genus in 1999 een monografische behandeling, waarbij aspecten van de anatomie en taxonomie, en de functies van de kam aan de orde kwamen. Williamson publiceerde later een onafhankelijke review van de resten, waarin hij het niet eens was met de taxonomische conclusies.
John Ostrom beschreef een ander goed specimen (FMNH P27393) uit New Mexico als P. cyrtocristatus in 1961. Het omvat een gedeeltelijke schedel met een korte, afgeronde kuif, en een groot deel van het postcraniale skelet met uitzondering van de voeten, de nek en delen van de staart. De specifieke naam is afgeleid van het Latijnse curtus “verkort” en cristatus “gekuifd”. Het exemplaar werd gevonden in de top van de Fruitland Formatie of, waarschijnlijker, aan de basis van de bovenliggende Kirtland Formatie. Het verspreidingsgebied van deze soort werd uitgebreid in 1979, toen David B. Weishampel en James A. Jensen een gedeeltelijke schedel met een soortgelijke kuif (BYU 2467) beschreven uit de Kaiparowits Formatie van het Campanien in Garfield County, Utah. Sindsdien is in Utah nog een schedel gevonden met de korte/ronde P. cyrtocristatus-kam morfologie.
Soorten
Parasaurolophus walkeri schedelafgietsel, Natural History Museum, Londen.
Parasaurolophus cyrtocristatus schedelafgietsel, Field Museum of Natural History.
De typesoort P. walkeri, uit Alberta, is bekend van één enkel specimen. Hij verschilt van P. tubicen door eenvoudiger buizen in de kuif, en van P. cyrtocristatus door een lange, onronde kuif en een langere bovenarm dan onderarm.
P. tubicen, uit New Mexico, is bekend van de overblijfselen van ten minste drie individuen. Het is de grootste soort, met complexere luchtgangen in de kuif dan P. walkeri, en een langere, rechtere kuif dan P. cyrtocristatus.
P. cyrtocristatus, uit New Mexico en Utah, is bekend van drie mogelijke specimens. Het is de kleinste soort, met een korte afgeronde kuif. De geringe grootte en de vorm van de kuif hebben verscheidene wetenschappers ertoe gebracht te suggereren dat het juvenielen of wijfjes zijn van P. tubicen, die uit ongeveer dezelfde tijd en uit dezelfde formatie in New Mexico afkomstig is. Zoals opgemerkt door Thomas Williamson is het type-materiaal van P. cyrtocristatus ongeveer 72% van de grootte van P. tubicen, dicht bij de grootte waarbij andere lambeosauriërs volgens de interpretatie definitief sexueel dimorfisme in hun kuif beginnen te vertonen (~70% van volwassen grootte). Dit standpunt is verworpen in recente reviews van lambeosaurines.
Paleobiology
Paleoecology
Gorgosaurus (links) die een Parasaurolophus cyrtocristatus aanvalt.
Parasaurolophus walkeri, uit de Dinosaur Park Formatie, was lid van een diverse en goed gedocumenteerde fauna van prehistorische dieren, waaronder bekende dinosaurussen als de gehoornde Centrosaurus, Styracosaurus, en Chasmosaurus; mede-eendinosauriërs als Prosaurolophus, Gryposaurus, Corythosaurus en Lambeosaurus; de tyrannosauriër Gorgosaurus; en de gepantserde Edmontonia en Euoplocephalus. Hij was een zeldzaam bestanddeel van deze fauna. De Dinosaur Park Formatie wordt geïnterpreteerd als een laag-reliëf omgeving van rivieren en uiterwaarden die in de loop der tijd moerassiger werden en beïnvloed door mariene omstandigheden toen de Westelijke Binnenzee zich westwaarts verplaatste. Het klimaat was warmer dan in het huidige Alberta, zonder vorst, maar met nattere en drogere seizoenen. Coniferen waren kennelijk de dominante kroonplanten, met een ondergroei van varens, boomvarens, en angiospermen.
De Nieuw-Mexicaanse soorten deelden hun omgeving met de grote sauropode Alamosaurus, de eendenbek Kritosaurus, de gehoornde Pentaceratops, de gepantserde Nodocephalosaurus, de Saurornitholestes, en de nu nog naamloze tyrannosauriden. De Kirtland Formatie wordt geïnterpreteerd als riviervloedvlaktes die ontstonden na het terugtrekken van de Westelijke Binnenzee. Coniferen waren de dominante planten, en chasmosaurine gehoornde dinosauriërs kwamen blijkbaar vaker voor dan hadrosauriden.
Voeding
Als hadrosauride was Parasaurolophus een grote herbivoor op twee of vier poten, die planten at met een geraffineerde schedel die een slijpbeweging toeliet analoog aan kauwen. Zijn tanden werden voortdurend vervangen en samengebald in tandbatterijen die honderden tanden bevatten, waarvan er slechts een handvol tegelijk in gebruik waren. Hij gebruikte zijn snavel om plantaardig materiaal te oogsten, dat in de kaken werd gehouden door een wangachtig orgaan. Het eten vond plaats vanaf de grond tot ongeveer 4 meter daarboven. Zoals opgemerkt door Bob Bakker, hebben lambeosaurines smallere snavels dan hadrosaurines, hetgeen impliceert dat Parasaurolophus en zijn verwanten zich selectiever konden voeden dan hun breedgesnavelde, kuifloze soortgenoten.
Craniale kuif
Vele hypothesen zijn naar voren gebracht over de functies die de craniale kuif van Parasaurolophus vervulde, maar de meeste zijn in diskrediet gebracht. Thans wordt aangenomen dat hij verschillende functies kan hebben gehad: visueel vertoon voor het identificeren van soort en geslacht, geluidsversterking voor communicatie, en thermoregulatie. Het is niet duidelijk welke functie het belangrijkst was op welk moment in de evolutie van de kam en de inwendige neusgangen.
Verschillen tussen soorten en groeistadia
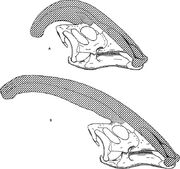
Diagram dat de nariakammen van Parasaurolophus cyrtocristatus (a) en Parasaurolophus walkeri (b) vergelijkt.
Zoals voor andere lambeosaurines wordt aangenomen, dat de schedelkam van Parasaurolophus veranderde met de leeftijd en een sexueel dimorf kenmerk was bij volwassenen. James Hopson, een van de eerste onderzoekers die lambeosaurinekammen beschreef in termen van een dergelijk onderscheid, suggereerde dat P. cyrtocristatus, met zijn kleine kuif, de vrouwelijke vorm was van P. tubicen. Thomas Williamson stelde voor dat het de juveniele vorm was. Geen van beide hypothesen werd algemeen aanvaard. Aangezien slechts zes goede schedels en één juveniele hersenpan bekend zijn, zal aanvullend materiaal deze mogelijke verwantschappen helpen ophelderen. Williamson merkte op dat in ieder geval de jonge Parasaurolophus waarschijnlijk kleine, afgeronde kammen had zoals P. cyrtocristatus, die waarschijnlijk sneller groeiden naarmate de individuen volwassener werden. Recent heronderzoek van een juveniele hersenkast die eerder aan Lambeosaurus werd toegewezen, en nu aan Parasaurolophus, levert het bewijs dat een kleine buisvormige kam aanwezig was bij juvenielen. Dit specimen bewaart een kleine opwaartse uitwaaiering van de voorhoofdsbeenderen die gelijkaardig was aan maar kleiner dan wat wordt gezien in volwassen specimens; bij volwassenen vormden de voorhoofdsbeenderen een platform dat de basis van de kam ondersteunde. Dit exemplaar geeft ook aan dat de groei van de kuif bij Parasaurolophus en het gezichtsprofiel van juveniele individuen verschilden van het Corythosaurus-Hypacrosaurus-Lambeosaurus-model, deels omdat de kuif van Parasaurolophus de dunne benige ‘coxkam’ mist die het bovenste deel van de kuif van de andere drie lambeosaurines uitmaakt.
Verworpen hypothesen over functie
Vele vroege suggesties richtten zich op aanpassingen voor een aquatische levensstijl, in navolging van de hypothese dat hadrosauriden amfibisch waren, een gangbare gedachtegang tot in de zestiger jaren van de vorige eeuw. Zo stelde Alfred Sherwood Romer voor dat het diende als snorkel, Martin Wilfarth dat het een hulpstuk was voor een mobiele proboscis die werd gebruikt als adembuis of voor het verzamelen van voedsel, Charles M. Sternberg dat het diende als luchtvanger om water uit de longen te houden, en Ned Colbert dat het diende als luchtreservoir voor langdurig verblijf onder water.
Andere voorstellen waren meer natuurkundig van aard. Zoals hierboven vermeld, stelde William Parks voor dat het met ligamenten of spieren aan de wervels was verbonden, en hielp bij het bewegen en ondersteunen van het hoofd. Othenio Abel stelde voor dat het gebruikt werd als wapen in de strijd tussen leden van dezelfde soort, en Andrew Milner stelde voor dat het gebruikt zou kunnen worden als afweermiddel tegen gebladerte, zoals de helmkam (een “casque” genoemd) van de kasuaris. Weer andere voorstellen maakten de huisvesting van gespecialiseerde organen tot de belangrijkste functie. Halszka Osmólska suggereerde dat het zoutklieren huisvestte, en John Ostrom suggereerde dat het uitgebreide gebieden voor reukweefsel en veel verbeterde reukzin van de lambeosaurines huisvestte, die geen duidelijke verdedigingsmogelijkheden hadden. Een ongebruikelijke suggestie, gedaan door creationist Duane Gish, is dat de kam chemische klieren huisvestte die hem in staat stelden stralen chemisch “vuur” naar vijanden te werpen, vergelijkbaar met de hedendaagse bombardeerkever.
De meeste van deze hypotheses zijn in diskrediet gebracht of verworpen. Er is bijvoorbeeld geen gat aan het eind van de kuif voor een snorkelfunctie. Er zijn geen spierlittekens voor een proboscis en het is dubieus dat een dier met een snavel er een nodig zou hebben. Als luchtsluis zou het geen water hebben tegengehouden. Het voorgestelde luchtreservoir zou ontoereikend zijn geweest voor een dier van de grootte van Parasaurolophus. Andere hadrosauriden hadden grote koppen zonder dat zij grote holle kammen nodig hadden om als bevestigingspunten voor ondersteunende ligamenten te dienen. Ook verklaart geen van de voorstellen waarom de kam zo’n vorm heeft, waarom andere lambeosauriërs kammen zouden moeten hebben die er heel anders uitzien maar een soortgelijke functie vervullen, hoe kuifloze of hadrosauriërs met vaste kammen zich zonder dergelijke mogelijkheden konden redden, of waarom sommige hadrosauriërs vaste kammen hadden. Deze overwegingen zijn vooral van invloed op hypotheses die gebaseerd zijn op het vergroten van de mogelijkheden van systemen die al in het dier aanwezig zijn, zoals de hypothese van de zoutklier en die van de olfactor, en wijzen erop dat dit niet de primaire functies van de kam waren. Bovendien blijkt uit werk aan de neusholte van lambeosaurines dat reukzenuwen en bijbehorend zintuiglijk weefsel zich grotendeels buiten het deel van de neusgangen in de kam bevonden, zodat de uitbreiding van de kam weinig te maken had met de reukzin.
Sociale functies
Levensherstel van Parasaurolophus walkeri met hypothetische huidrand en strepen op de kuif.
In plaats daarvan hebben sociale en fysiologische functies meer steun gekregen als functie(s) van de kuif, waarbij de nadruk ligt op visuele en auditieve identificatie en communicatie. Als groot object heeft de kuif een duidelijke waarde als visueel signaal, en onderscheidt dit dier zich van zijn tijdgenoten. De grote afmetingen van de oogkassen van hadrosauriden en de aanwezigheid van sclerotische ringen in de ogen impliceren een scherp gezichtsvermogen en dagelijke gewoonten, bewijs dat het gezichtsvermogen belangrijk was voor deze dieren. Als, zoals gewoonlijk wordt geïllustreerd, een huidrand zich uitstrekte van de kam naar de nek of rug, zou de voorgestelde visuele vertoning veel opzichtiger zijn geweest. Zoals door andere schedels van lambeosauriërs wordt gesuggereerd, maakte de kam van Parasaurolophus waarschijnlijk zowel soortidentificatie mogelijk (zoals het onderscheiden van Corythosaurus of Lambeosaurus) als determinatie tussen mannetjes en vrouwtjes, gebaseerd op vorm en grootte.
Sounding function
Computermodel van de neusdoorgangen in de kam van P. tubicen.
Het uiterlijke uiterlijk van de kam komt echter niet overeen met de complexe interne anatomie van de neusdoorgangen, wat een andere functie suggereert die het gebruik van de interne ruimte verklaart. Carl Wiman was de eerste die, in 1931, voorstelde dat de doorgangen een auditieve signaalfunctie hadden, zoals een trommelhoorn; Hopson en David B. Weishampel hebben dit idee in de jaren 1970 en 1980 opnieuw bekeken. Hopson vond dat er anatomisch bewijs is dat hadrosauriden een sterk gehoor hadden. Er is tenminste één voorbeeld, bij de verwante Corythosaurus, van een slanke stapes (reptielachtig oorbeen) op zijn plaats, hetgeen in combinatie met een grote ruimte voor een trommelvlies een gevoelig middenoor impliceert. Bovendien is de laga van de hadrosauride langgerekt als die van een krokodil, wat erop wijst dat het auditieve deel van het binnenoor goed ontwikkeld was. Weishampel suggereerde dat P. walkeri in staat was frequenties van 48 tot 240 Hz te produceren, en P. cyrtocristatus (geïnterpreteerd als een juveniele kuifvorm) 75 tot 375 Hz. Gebaseerd op overeenkomsten tussen de binnenoren van hadrosauriden en die van krokodillen, stelde hij ook voor dat volwassen hadrosauriden gevoelig waren voor hoge frequenties, zoals hun nageslacht die zou kunnen produceren. Volgens Weishampel is dit in overeenstemming met de mogelijkheid dat ouders en nakomelingen met elkaar communiceren.
Computer modellering van een goed bewaard gebleven exemplaar van P. tubicen, met complexere luchtwegen dan die van P. walkeri, heeft de reconstructie mogelijk gemaakt van het mogelijke geluid dat zijn kuif produceerde. Het hoofdpad resoneert bij ongeveer 30 Hz, maar de gecompliceerde anatomie van de sinus veroorzaakt pieken en dalen in het geluid.
Koelingsfunctie
Het grote oppervlak en de vascularisatie van de kam suggereren ook een thermoregulerende functie. P.E. Wheeler stelde dit gebruik in 1978 voor het eerst voor als een manier om de hersenen koel te houden. Teresa Maryańska en Osmólska stelden rond diezelfde tijd ook thermoregulatie voor, en Sullivan en Williamson namen verdere belangstelling op. David Evans’ 2006 bespreking van lambeosaurine kuif functies was gunstig voor het idee, althans als een initiële factor voor de evolutie van kuif uitbreiding.
In De Media
Parasaurolophus is een zeer populaire dinosaurus. Het was in de film Disney’s Dinosaur als een kudde lid. Hij kwam ook meerdere malen voor in de beroemde documentaire Clash of the Dinosaurs. Hij verscheen ook in het populaire TV-programma Prehistoric Park, waar hij de prooi werd van de grote carnivoren Deinosuchus en Albertosaurus. Hij verscheen een paar keer in de Jurassic Park-films, als lid van de kudde in de eerste film, vervolgens gevangen gehouden door jagers in de tweede en meelopend met Corythosaurus in de derde. en het personage Dweeb in “We’re Back! A Dinosaur’s Story,” is zelf een Parasaurolophus.
Parasaurolophus komt ook voor in Turok, als een volgzame planteneter die normaal gesproken niet schadelijk is, maar agressief kan zijn als hij ernstig wordt geprovoceerd.
Er is ook een Parasaurolophus zord in Power Rangers: Dino Thunder en Power Rangers: Dino Charge.
Parasaurolophus komt kort voor aan het begin van de Disney Pixar film The Good Dinosaur.
Zie ook
Parasaurolophus/Gallery
- Lull, Richard Swann Wright, Nelda E. Hadrosaurian Dinosaurs of North America, page 229. Gepubliceerd: 1942, Geological Society of America, Geological Society of America Special Paper 40
- 2.0 2.1 2.2 2.3 Glut, Donald F. Dinosaurs: The Encyclopedia, Chapter: Parasaurolophus, pagina’s 678-684. Gepubliceerd: 1997, McFarland & Co, in Jefferson, North Carolina. ISBN 0-89950-917-7
- 3.0 3.1 3.2 Lull and Wright, Hadrosaurian Dinosaurs of North America, pp. 209-213.
- Brett-Surman, Michael K. en Wagner, Jonathan R. Carpenter, Kenneth (ed.) Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, hoofdstuk: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids, blz. 135-169. Gepubliceerd, 2006, Indiana University Press, in Bloomington en Indianapolis ISBN 0-253-34817-X
- 5.0 5.1 5.2 5.3 5.4 5.5 Horner, John R., Weishampel, David B.; en Forster, Catherine A, Weishampel, David B.; Osmólska, Halszka; en Dodson, Peter (eds.) The Dinosauria, 2nd edition, chapter: Hadrosauridae, blz. 438-463. Gepubliceerd: 2004, University of California Press, in Berkeley. ISBN 0-520-24209-2
- 6.0 6.1 6.2 6.3 6.4 6.5 Parks, William A. Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, deel 13, pagina’s 1-32. Gepubliceerd: 1922, University of Toronto Studies, Geology Series.
- 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 Sullivan, Robert M. and Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, uit de serie New Mexico Museum of Natural History and Science Bulletin, 15, pages 1-52. Gepubliceerd: 1999, New Mexico Museum of Natural History and Science, in Albuqueque, New Mexico.
- 8.0 8.1 8.2 Ostrom, John H. 1961 A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, Journal of Paleontology, Volume 35, 3rd issue, on pages 575-577.
- Gilmore, Charles W., On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, volume 38, issue 43, pages 29-48, Parks. Gepubliceerd: 1924, Canada Department of Mines Geological Survey Bulletin (Geological Series)
- Weishampel, David B. en Horner, Jack R., Weishampel, David B.; Osmólska, Halszka; en Dodson, Peter (eds.) The Dinosauria, 1e editie, Hoofdstuk: Hadrosauridae, blz. 534-561. Gepubliceerd: 1990, University of California Press in Berkeley. ISBN 0-520-06727-4
- 11.0 11.1 11.2 11.3 Evans, David C., and Reisz, Robert R. 2007. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaurid (Ornithischia) from the Dinosaur Park Formation, Alberta, uit het Journal of Vertebrate Paleontology, vol. 27 issue 2, op pagina’s 373-393.
- Godefroit, Pascal, Shuqin Zan; and Liyong Jin. 2000. Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaurine from the Late Maastrichtian of northeastern China, uit de Compte Rendus de l’Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, pages 875-882.
- 13.0 13.1 13.2 13.3 13.4 Williamson, Thomas E. 2000. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico Lucas, S.G.; and Heckert, A.B. (eds.) Dinosaurs of New Mexico, from the New Mexico Museum of Natural History and Science Bulletin, 17 Uitgegeven door het New Mexico Museum of Natural History and Science, in Albuqueque, New Mexico. Pagina’s 191-213.
- Liddell, Henry George en Robert Scott, 1980. A Greek-English Lexicon (Verkorte uitgave) Uitgegeven: Oxford University Press in het Verenigd Koninkrijk. ISBN 0-19-910207-4
- 15.0 15.1 Ryan, Michael J. and Evans, David C., Currie, Phillip J., and Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, Hoofdstuk: Ornithischian Dinosaurs. 2005, Gepubliceerd: Indiana University Press, in Bloomington. Pagina’s 312-348, ISBN 0-253-34595-2
- 16.0 16.1 Evans, David C., Reisz, Robert R.; and Dupuis, Kevin, 2007. A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus. Journal of Vertebrate Paleontology, vol. 27, issue 3 pages 642-650.
- Currie, Phillip J; and Russell, Dale A. “Geographic and stratigraphic distribution of dinosaur remains” in Dinosaur Provincial Park, p. 553.
- Weishampel, David B. (1990). “Dinosaurus Distributie”, in The Dinosauria (1st), pp. 63-139.
- 19.0 19.1 19.2 Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). “Dinosauria Distribution”, in The Dinosauria (2nd), pp. 517-606.
- 20.0 20.1 Wiman, Carl, 1931. Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico, uit de Nova Acta Regia Societas Scientarum Upsaliensis, serie 4, vol. 7, issue 5. (Duits). Pagina’s 1-11.
- 21.0 21.1 Simpson, D.P. Cassell’s Latin Dictionary, uitgegeven door Cassell Ltd. 1979, editie 5, in Londen. ISBN 0-304-52257-0 Blz. 883.
- Weishampel, David B. en Jensen, James A. 1979. Parasaurolophus (Reptilia: Hadrosauridae) uit Utah, uit het Journal of Paleontology, vol. 53, issue 6, pages 1422-1427.
- Eberth, David A. 2005. “De geologie”, in Dinosaur Provincial Park, blz. 54-82.
- Braman, Dennis R., en Koppelhus, Eva B. 2005. “Campanian palynomorphs”, in Dinosaur Provincial Park, pp. 101-130.
- Russell, Dale A. An Odyssey in Time: Dinosaurussen van Noord-Amerika, 1989. Uitgever: NorthWord Press, in Minocqua, Wisconsin. ISBN 1-55971-038-1 Pagina’s 160-164.
- Bakker, Robert T. 1986. De dinosaurusketterijen: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, uitgegeven door William Morrow, in New York. ISBN 0-8217-2859-8 Pagina 194.
- 27.0 27.1 27.2 Norman David B. The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom, hoofdstuk: Hadrosaurids II. 1985. Uitgegeven door Crescent Books, in New York. Pagina’s 122-127. ISBN 0-517-468905
- 28.0 28.1 28.2 28.3 28.4 Evans, David C., 2006. Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs, uit het Journal of Paleobiology, vol. 32, issue 1, Pages 109-125.
- 29.0 29.1 29.2 29.3 Hopson, James A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs , uit het Journal of Paleobiology, volume one, issue one, pages 21-43.
- Romer. Alfred Sherwood, 1933. Vertebrate Paleontology, van de University of Chicago Press, op pagina 491.
- Wilfarth, Martin, 1947. Russeltragende Dinosaurier, uit de Journal of Orion (München), vol. 2. pp. 525-532 (Duitse taal).
- Sternberg, Charles M. 1935. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous from the Journal of the Canada Department of Mines Bulletin (Geological Series), volume 77, issue 52, on pages 1-37.
- Colbert, Edwin H. The Dinosaur Book: The Ruling Reptiles and their Relatives, in 1945 uitgegeven door het American Museum of Natural History, Man and Nature Publications, 14, in New York. Pagina 156.
- Abel, Othenio, 1924. Die neuen Dinosaurierfunde in der Oberkreide Canadas uit het tijdschrift Jarbuch Naturwissenschaften, jaargang 12, nummer 36, op pagina’s 709-716. (Duits) 1924.
- Osmólska, Halszka, 1979. Nasale zoutklieren bij dinosauriërs, uit het tijdschrift Acta Palaeontologica Polonica, vol. 24, blz. 205-215.
- 36.0 36.1 Ostrom, John H., 1962. The cranial crests of hadrosaurian dinosaurs, uit het Tijdschrift voor Postilla, vol. 62, pp. 1-29.
- Gish, Duane T., 1992. Dinosaurs by Design, uitgegeven door Master Books, in Green Forest. ISBN 0-89051-165-9 Pagina 82.
- 38.0 38.1 Weishampel, David B., 1981. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), uit het Tijdschrift voor Paleobiologie, vol. 7, issue 2, pp. 252-261.
- Weishampel David B., 1981. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies, uit het Tijdschrift voor Paleontologie, vol. 55, issue 5, pp. 1046-1057.
- Wetenschappers gebruiken digitale paleontologie om stem van Parasaurolophus Dinosaurus te produceren, door de Sandia National Laboratories (1997-12-05). Ontvangen op 20 januari 2009.
- Diegert, Carl F. en Williamson, Thomas E., 1998. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen uit het Journal of Vertebrate Paleontology, vol. 18, issue 3, Suppl. p. 38A.
- Sullivan, Robert M. and Williamson, Thomas E., 1996. Een nieuwe schedel van Parasaurolophus (langkuifvorm) uit New Mexico: uiterlijke en inwendige (CT-scans) kenmerken en hun functionele implicaties, uit het Journal of Vertebrate Paleontology, vol. 16, issue 3, Suppl. pp.68A.
- Wheeler, P.E., 1978. Elaborate CNS cooling structure in large dinosaurs Journal of Nature, vol. 275, on pp. 441-443.
- Maryańska, Teresa en Osmólska, Halszka, in 1979. Aspecten van de schedelanatomie van hadrosauriërs, uit het Tijdschrift voor Lethaia, vol. 12, op pp. 265-273.
- Wetenschappers gebruiken Digitale Paleontologie om Stem van Parasaurolophus Dinosaurus te produceren; van Sandia National Laboratories.
- Restauraties van P. walkeri en P. cyrtocristatus, op dezelfde schaal, door Scott Hartman; bij Skeletal Drawing.com.
- Parasaurolophus pagina op Enchanted Learning.com