
Grafik anvendt med tilladelse. Daniel D. Brown, LaughingMantis.com.
Echinodermer er en af de mest højt afledte dyregrupper med mange arter som vigtige komponenter i flere marine samfund. De er klassificeret efter tre grundlæggende fælles karakteristika: 1) pentaradial symmetri, 2) skelet lavet af tredimensionelle kalcitiske elementer og 3) tilstedeværelsen af et vandkarsystem med en ekstern åbning. Især dette vand-karsystem (WVS) er fortsat en unik anvendelse af et hydraulisk system, der genererer trykgradienter, der er i stand til at kommandere hundredvis af små, bedårende rørfødder til at kravle over ujævne overflader og ernære sig af hårdføre byttedyr med skal som muslinger.
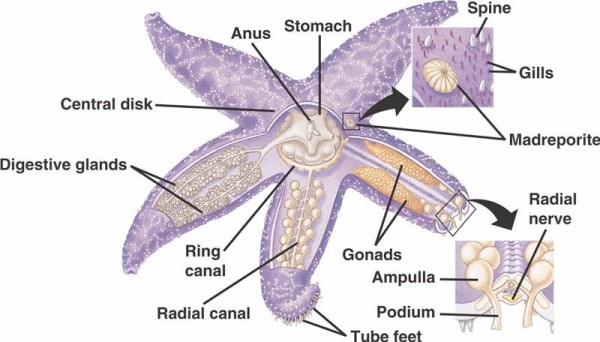
De grundlæggende komponenter i WVS hos ikke-crinoide pighuder (nedenfor) omfatter en circumoral ringkanal med fem radiale kanaler, der strækker sig ud fra den. Til ringkanalen er også knyttet madreporitten, der fungerer som en dør mellem havets ydre miljø og pighuderens indre miljø. Madreporitten, der er opkaldt efter sin lighed med korallen Madreporaria (der betyder “porernes mor”), er foret med celler, der slår svanslignende flageller i forskellige retninger. væsken inde i WVS’en ligner havvand i sammensætning, men indeholder op til 60 % flere kaliumioner, lidt højere koncentrationer af kloridioner og frit flydende albuminholdigt protein. Man har også fundet, at søagurker har røde blodlegemer i deres WVS.
Hver radialkanal har serielt gentagne par af kortere kanaler, der er fastgjort til rørfødderne, som betjenes ved muskelkontraktion af de ampuller, der er placeret direkte over dem. Tænk på en kalkunpære: Når du klemmer på pæren, skubber den væske ud i pæreenden, slipper du trykket, suger den væske ind mod pæren igen. Crinoider adskiller sig fra de andre, fordi de mangler både disse kortere kanaler og ampullerne, og deres rørfødder forekommer i grupper af tre, der kommer direkte fra en kanal, der ligner radialkanalen.
Crinoiderne bevarer den mest primitive form af WVS. Man mente, at rørfødder oprindeligt blev brugt udelukkende til åndedræt samt som fødesøgningsstrukturer, men fossile eocrinoider har tydelige porer mellem skeletpladerne, hvilket fik Nichols (1972) til at konkludere, at rørfodssystemet ikke var særlig omfattende tidligt i crinoidernes udvikling. Men han tager ikke hensyn til andre fortolkninger af strukturerne, såsom åbninger til indre dele, der er relateret til lokomotion eller fødeindtagelse og ikke har noget med respiration at gøre. Karpoider, såsom Gyrocystis og Dendrocystites, der er fundet i sedimenter fra det mellemste Kambrium, anses for at være basale i forhold til pighuderlinjen på grund af lighed i skeletelementer og antages at have bløde, fremtrækkelige indre tentakulære strukturer. Eksistensen af en primitiv WVS er ikke entydig i de fossile optegnelser af carpoider, men den anses for at have udviklet sig forud for andre pighuderkarakteristika. Den tidligste antydning af noget, der ligner en WVS, stammer fra Helicoplacoidea, der er fundet i nedre kambriske aflejringer (bemærket ved tilstedeværelsen af stærkt organiserede rækker af ambulacrale porer). Ifølge Paul (1977) har ingen moderne klasse divergeret fra carpoiderne, som har været uddøde siden Mesozoikum. Helicoplacoiderne, med triradial symmetri, divergerede meget tidligt fra carpoiderne, og resten af pighuderlinjen udviklede sig på Helicoplacoid bauplan.

Xyloplax fra Voight 2005.
Selv om pighudernes stamtræ ikke er helt gennemarbejdet, oplevede de en kort diversifikationsbegivenhed i løbet af kambrisk og ordivicium, og den grundlæggende bauplan er siden da forblevet lidt ændret. De fire ikke-crinoide klasser har lignende struktur og funktion i deres WVS. Holothuroiderne er sandsynligvis de mest afledte på klasseniveau, fordi deres bucchale fodringstentakler er forlænget fra den circumorale ringkanal og ikke indeholder nogen skeletelementer. Den nyligt opdagede Xyloplax (til venstre), en mærkelig slags søstjerne, har et mere unikt WVS-arrangement: et nyt system med dobbelte ringkanaler med interradiale forbindelser og et monoserielt sæt af rørfødder. Janies og McEdward (1994) foreslår, at den unikke geometri af dens WVS er opstået via en ændring i udviklingsmekanismen hos en juvenil velatid Asteroid. Dette tilfælde repræsenterer den mest moderne og afledte udvikling af WVS.
Stikpindsvinenes hurtige diversificering, manglen på god bevaring og fraværet af mere end ét typeeksemplar til sammenligning af obskure fossilklasser udgør problemer, når man forsøger at forklare deres udviklingshistorie. WVS er strengt taget ikke kun et unikt pighuderkarakteristikum. Der er ligheder med strukturer hos hemichordaterne og den hydraulisk drevne lophophore. For ikke at nævne, at embryologiske karakteristika hos pighuder også deles blandt chordater, hemichordater og lophophorater. Desuden udviklede ringkanalen sig fra en U-formet forløber, der til sidst smelter sammen i løbet af ontogenesen for at danne den ring, som vi ser hos pighuder i dag. Dette synes at passe med Nichols’ (1967, 1972) teori om, at pighuder delte forfædre med en lophophorate-lignende forgænger, som også har U-formede tarme i både larve- og voksenformer.
WVS er en unik og, i nutidens pighuder, vellykket innovation. Det virker plausibelt, at det indviklede netværk af kanaler kunne være bygget på allerede eksisterende anatomiske strukturer, såsom lophophoren og den U-formede tarm hos en lophophatlignende fælles forfader, men beviserne er uklare. Det, der er slående for mig, er, at en sådan nyskabelse ikke findes i andre stamceller. WVS’en opstod sandsynligvis som en fødestruktur hos tidlige sessile pighuder, som det fremgår af fossile fortolkninger, og blev sekundært brugt til at bevæge sig. En anden fortolkning kan være, at WVS udviklede sig mere end én gang blandt uddøde og uddøde pighuder til forskellige formål (dvs. åndedræt, fødeindtagelse, fremdrift). Uanset hvad er WVS en forbløffende tilpasning blandt echinodermata.

Fra Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) A new class of Echinodermata from New Zealand. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) The water vascular system and functional morphology of Paleozoic asteroids. Lethaia 21: 189-206.
3) Binyon J. (1972) Physiology of Echinoderms. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) A re-appraisal of the fluid loss resulting from operation of the water vascular system of the starfish of the starfish, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrates. Sinauer Associates, Inc. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: Vol. IV Echinodermata. McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) A hypothesis for the evolution of the Concentricycloid water-vascular system. I Wilson Jr. W.H., Stricker S.A., Stricker S.A., Shinn G.L. (red.): Reproduction and Development of Marine Invertebrates. John Hopkins University Press, Baltimore.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-some implications of the sister-group relationship between echinoderms and chordates. I Paul C.R.C., Smith A.B. (eds.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, London: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) The early phylogeny of chordates and echinoderms and the origin of chordate left-right symmetry and bilateral symmetry. Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms. Croom Helm Ltd., London.
13) Lowe C.J., Wray G.A. (1997) Radikale ændringer i homeobox-genernes roller i løbet af pighudernes evolution. Nature 389: 718-721.
14) Menge B.A. (1982) Effekter af fødeindtagelse på miljøet: Asteroidea. I Jangoux M., Lawrence J.M. (eds.): Echinoderm Nutrition. A.A. Balkema, Rotterdam: 521-552.
15) Nichols D. (1967) The origin of echinoderms. In Millott N. (ed.): Echinoderm Biology. Academic Press Inc., London: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontology 15(4): 519-538.
17) Paul C.R.C. (1977) Evolution af primitive pighuder. In Hallam A. (ed.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Comments on the water vascular system, food grooves, and ancestry of Clypeasteroid Echinoids. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) Vedligeholdelse af væskevolumen i vandkarsystemet hos søstjerner. Nature 262: 577-578.
21) Smith A.B. (1988) To group or not to group: Den taksonomiske position af Xyloplax. I Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (eds.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Some aspects of evolution in Echinodermata. In de Beer O.R. (ed.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, London: 287-303.
23) Strathmann R.R. (1975) Limitations on diversity of forms: Branching of ambulacral systems of echinoderms. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific (Første rapport om den gådefulde pighvar Xyloplax fra det nordlige Stillehav). Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: Funktioner og molekylære tilpasninger. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Functional bauplan of the echinoderms. I Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) Genes involved in the evolution of echinoderm morphology. In Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 232.
Share the post “Veins of Water: The Evolution of the Echinoderm Water-Vascular System”
- Shares
 Kevin Zelnio (870 Posts)
Kevin Zelnio (870 Posts)