
Grafico usato con permesso. Daniel D. Brown, LaughingMantis.com.
Gli echinodermi sono uno dei gruppi di animali più altamente derivati con molte specie come componenti significativi di diverse comunità marine. Sono classificati da tre caratteristiche fondamentali condivise: 1) simmetria pentaradiale, 2) scheletro fatto di elementi calcitici tridimensionali, e 3) la presenza di un sistema idrovascolare con un’apertura esterna. Questo sistema idrico-vascolare (WVS), in particolare, rimane un’applicazione unica di un sistema idraulico, generando gradienti di pressione che sono in grado di comandare centinaia di minuscoli, adorabili piedi tubolari per strisciare su superfici irregolari e nutrirsi di dure prede sgusciate come le cozze.
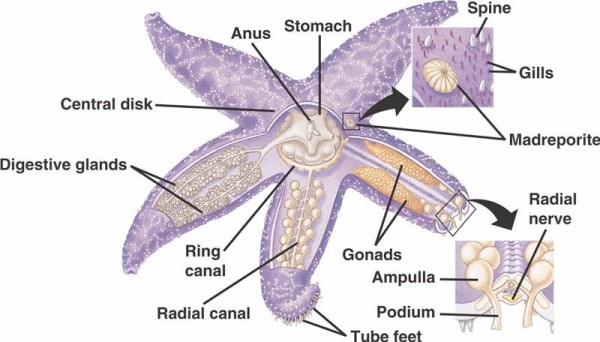
I componenti fondamentali del WVS negli echinodermi non-crinoidi (sotto) includono un canale anulare circumorale con cinque canali radiali che si estendono da esso. Al canale anulare è attaccato anche il madreporite, che funge da porta tra l’ambiente esterno del mare e l’ambiente interno dell’echinoderma. Il madreporite, chiamato così per la sua somiglianza con il corallo Madreporaria (che significa “madre dei pori”), è foderato con cellule che battono flagelli simili a code in una varietà di direzioni. Il fluido all’interno del WVS è simile all’acqua di mare nella composizione, ma contiene fino al 60% in più di ioni di potassio, concentrazioni leggermente superiori di ioni cloruro, e proteine albuminose liberamente fluttuanti. I cetrioli di mare hanno anche trovato dei globuli rossi nel loro WVS.
Ogni canale radiale ha coppie di canali più corti che si ripetono in serie e si attaccano ai piedi del tubo, che sono azionati dalla contrazione muscolare delle ampolle situate direttamente sopra di loro. Pensate alla pipetta da tacchino: quando schiacciate il bulbo, esso spinge il fluido fuori dall’estremità della pipetta, lasciate andare la pressione e il fluido viene risucchiato verso il bulbo. I crinoidi differiscono perché mancano sia di questi canali più corti che delle ampolle e i loro piedi tubolari si presentano in gruppi di tre che provengono direttamente da un canale simile al canale radiale.
I crinoidi conservano la forma più primitiva del WVS. Si credeva che i piedi a tubo fossero stati originariamente usati interamente per la respirazione e per l’alimentazione, ma gli eocrinoidi fossili hanno pori distinti tra le piastre scheletriche che hanno portato Nichols (1972) a concludere che il sistema dei piedi a tubo non era molto esteso all’inizio dell’evoluzione dei crinoidi. Ma egli non prende in considerazione altre interpretazioni delle strutture, come aperture per parti interne legate alla locomozione o all’alimentazione e non collegate alla respirazione. I carpoidi, come Gyrocystis e Dendrocystites, trovati nei sedimenti del Cambriano medio sono considerati basali alla stirpe degli echinodermi per la somiglianza degli elementi scheletrici e si ipotizza che abbiano strutture tentacolari interne morbide e protrattili. L’esistenza di un WVS primitivo è inconcludente nel record fossile dei carpoidi, ma si ritiene che si sia evoluto prima di altre caratteristiche degli echinodermi. Il primo suggerimento di qualcosa di simile a un WVS proviene dagli Helicoplacoidea trovati nei depositi del Cambriano inferiore (noti per la presenza di file altamente organizzate di pori ambulacrali). Secondo Paul (1977) nessuna classe moderna si è differenziata dai carpoidi, che si sono estinti dal Mesozoico. Gli elicoplacoidi, con simmetria triradiale, si sono differenziati molto presto dai carpoidi e il resto della stirpe degli echinodermi si è evoluto sul bauplan degli elicoplacoidi.

Xyloplax da Voight 2005.
Sebbene l’albero genealogico degli echinodermi non sia del tutto definito, essi hanno vissuto un breve evento di diversificazione durante il Cambriano e l’Ordiviciano e il bauplan di base è rimasto da allora poco cambiato. Le quattro classi non-crinoidi hanno una struttura e una funzione simili nel loro WVS. Gli oloturoidi sono probabilmente i più derivati a livello di classe, perché i loro tentacoli di alimentazione buccali sono estesi dal canale anulare circumorale e non contengono elementi scheletrici. Lo Xyloplax (a sinistra), una strana specie di stella di mare, scoperto di recente, ha una disposizione WVS più unica: un nuovo sistema con doppi canali anulari che contengono connessioni inter-radiali e una serie monoseriale di piedi a tubo. Janies e McEdward (1994) suggeriscono che la geometria unica del suo WVS si è evoluta attraverso una modifica nel meccanismo di sviluppo di un Asteroide velatide giovanile. Questo caso rappresenta l’evoluzione più moderna e derivata del WVS.
La rapida diversificazione degli echinodermi, la mancanza di una buona conservazione e l’assenza di più di un esemplare tipo per confrontare oscure classi fossili presenta problemi quando si cerca di spiegare la loro storia evolutiva. Il WVS non è, in senso stretto, solo una caratteristica unica degli echinodermi. Ci sono somiglianze con le strutture degli emicordati e del liofori a funzionamento idraulico. Senza contare che le caratteristiche embriologiche degli echinodermi sono condivise anche tra i cordati, gli emicordati e i liofori. Inoltre, il canale anulare si è sviluppato da un precursore a forma di U che alla fine si fonde insieme durante l’ontogenesi per formare l’anello che vediamo oggi negli echinodermi. Questo sembra combaciare con la teoria di Nichols (1967, 1972) che gli echinodermi hanno condiviso un’ascendenza con un predecessore simile ai liofori, che hanno anche viscere a forma di U sia nella forma larvale che in quella adulta.
Il WVS è un’innovazione unica e, negli echinodermi di oggi, di successo. Sembra plausibile che l’intricata rete di canali possa essere stata costruita su strutture anatomiche preesistenti, come il lioforo e l’intestino a forma di U di un antenato comune liofilo, ma le prove non sono chiare. Ciò che mi colpisce è che tale innovazione non è presente in altri phyla. Il WVS probabilmente è sorto come una struttura di alimentazione nei primi echinodermi sessili, come evidenziato dalle interpretazioni fossili, e secondariamente è stato utilizzato per la locomozione. Un’altra interpretazione può essere che il WVS si sia evoluto più di una volta tra gli echinodermi estinti ed esistenti per scopi diversi (cioè respirazione, alimentazione, locomozione). In ogni caso, il WVS è un adattamento sorprendente tra gli Echinodermata.

Da Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) Una nuova classe di Echinodermata dalla Nuova Zelanda. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) The water vascular system and functional morphology of Paleozoic asteroids. Lethaia 21: 189-206.
3) Binyon J. (1972) Physiology of Echinoderms. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) Una rivalutazione della perdita di fluido risultante dal funzionamento del sistema vascolare dell’acqua della stella marina, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrati. Sinauer Associates, Inc. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea e alcune possibili implicazioni. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: Vol. IV Echinodermata. McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Morfogeni celomici e idrovascolari altamente derivati in una stella marina con sviluppo pelagico diretto. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) Un’ipotesi per l’evoluzione del sistema idro-vascolare Concentricycloid. In Wilson Jr. W.H., Stricker S.A., Shinn G.L. (eds.): Riproduzione e sviluppo degli invertebrati marini. John Hopkins University Press, Baltimora.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-some implications of the sister-group relationship between echinoderms and chordates. In Paul C.R.C., Smith A.B. (eds.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, Londra: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) La filogenesi iniziale dei cordati e degli echinodermi e l’origine della simmetria sinistra-destra dei cordati e della simmetria bilaterale. Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms. Croom Helm Ltd., London.
13) Lowe C.J., Wray G.A. (1997) Alterazioni radicali nei ruoli dei geni homeobox durante l’evoluzione degli echinodermi. Nature 389: 718-721.
14) Menge B.A. (1982) Effetti dell’alimentazione sull’ambiente: Asteroidea. In Jangoux M., Lawrence J.M. (eds.): Echinoderm Nutrition. A.A. Balkema, Rotterdam: 521-552.
15) Nichols D. (1967) L’origine degli echinodermi. In Millott N. (ed.): Echinoderm Biology. Academic Press Inc., London: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontologia 15(4): 519-538.
17) Paul C.R.C. (1977) Evoluzione degli echinodermi primitivi. In Hallam A. (ed.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Commenti sul sistema vascolare acquatico, sui solchi alimentari e sull’ascendenza degli Echinoidi Clypeasteroidi. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) Mantenimento del volume del fluido nel sistema vascolare acquatico delle stelle marine. Nature 262: 577-578.
21) Smith A.B. (1988) Raggruppare o non raggruppare: La posizione tassonomica di Xyloplax. In Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (eds.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Alcuni aspetti dell’evoluzione in Echinodermata. In de Beer O.R. (ed.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, London: 287-303.
23) Strathmann R.R. (1975) Limitazioni alla diversità delle forme: Ramificazione dei sistemi ambulacrali degli echinodermi. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) La filogenesi dei deuterostomi e il gruppo gemello dei cordati: prove da molecole e morfologia. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Emoglobine nonvertebrate: Funzioni e adattamenti molecolari. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Bauplan funzionale degli echinodermi. In Candia Carnevali M.D., Bonasoro F. (eds.): Ricerca sugli echinodermi 1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) Geni coinvolti nell’evoluzione della morfologia degli echinodermi. In Candia Carnevali M.D., Bonasoro F. (eds.): Ricerca sugli echinodermi 1998. A.A. Balkema, Rotterdam: 232.
Condividi il post “Vene d’acqua: The Evolution of the Echinoderm Water-Vascular System”
- Shares
 Kevin Zelnio (870 Messaggi)
Kevin Zelnio (870 Messaggi)