
Gráfico usado com permissão. Daniel D. Brown, LaughingMantis.com.
Equinodermes são um dos grupos de animais mais altamente derivados com muitas espécies como componentes significativos de várias comunidades marinhas. Eles são classificados por três características fundamentais compartilhadas: 1) simetria pentaradial, 2) esqueleto feito de elementos calcíticos tridimensionais, e 3) presença de um sistema vascular aquático com uma abertura externa. Este sistema vascular de água (SVH), em particular, permanece uma aplicação única de um sistema hidráulico, gerando gradientes de pressão que são capazes de comandar centenas de pés de tubo minúsculos e adoráveis para rastejar sobre superfícies irregulares e se alimentar de presas com casca dura como mexilhões.
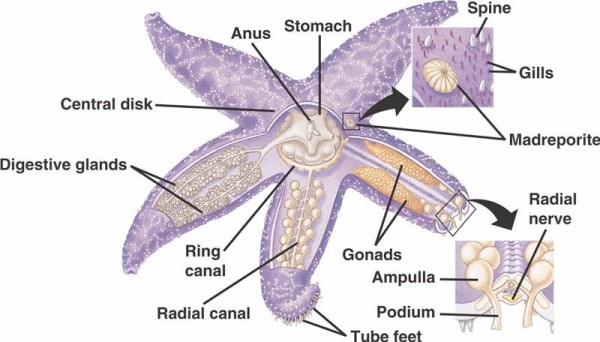
Os componentes básicos do SVH em equinodermos não crinoides (abaixo) incluem um canal circular circunoral com cinco canais radiais que se estendem de fora dele. Também ligado ao canal circular está o madreporite, que actua como uma porta entre o ambiente externo do mar e o ambiente interno do equinoderm. A madreporita, cujo nome deriva da sua semelhança com o coral Madreporaria (que significa “mãe dos poros”), é revestida com células que batem em flagelos de cauda em várias direcções. O fluido no interior do WVS é semelhante à água do mar em composição mas contém até 60% mais iões de potássio, concentrações ligeiramente superiores de iões cloreto e proteínas albuminosas que flutuam livremente. Os pepinos do mar também têm células vermelhas no seu WVS.
Cada canal radial tem repetidos em série pares de canais mais curtos que se ligam aos pés do tubo, que são operados pela contracção muscular das ampolas localizadas directamente sobre eles. Pense no peru baster: quando você aperta o bulbo empurra o fluido para fora da extremidade baster, solta a pressão e suga o fluido de volta para dentro em direção ao bulbo. Os crinóides diferem porque lhes faltam estes canais e ampolas mais curtos e os seus pés tubulares ocorrem em grupos de três vindos directamente de um canal semelhante ao canal radial.
Crinoides retêm a forma mais primitiva da SVB. Acreditava-se que os pés tubulares eram originalmente usados inteiramente para respiração, bem como estruturas de alimentação, mas os eocrinoides fósseis têm poros distintos entre placas esqueléticas que levaram Nichols (1972) a concluir que o sistema dos pés tubulares não era muito extenso no início da evolução dos crinoides. Mas ele não leva em consideração outras interpretações das estruturas, como aberturas para partes internas relacionadas à locomoção ou alimentação e não relacionadas à respiração. Carpoides, como Gyrocystis e Dendrocystites, encontrados em sedimentos do Cambriano médio são considerados basais à linhagem equinodérmica devido a similaridade em elementos esqueléticos e são hipotéticos de ter estruturas tendíneas internas macias e protraíveis. A existência de uma WVS primitiva é inconclusiva no registro fóssil de carpoides, mas considera-se que ela evoluiu antes de outras características equinodérmicas. A sugestão mais antiga de algo como uma WVS vem da Helicoplacoidea encontrada em depósitos cambrianos inferiores (notada pela presença de filas altamente organizadas de poros ambulacrais). Segundo Paul (1977), nenhuma classe moderna divergiu dos carpoides, que se extinguiram desde o Mesozóico. Helicoplacóides, com simetria triradial, divergiram muito cedo dos carpoides e o resto da linhagem equinodérmica evoluiu sobre o bauplan Helicoplacóide.

Xyloplax da Voight 2005.
Embora a árvore genealógica do equinoderm não esteja totalmente trabalhada, eles experimentaram um pequeno evento de diversificação durante o Cambriano e o Ordiviciano e o bauplan básico permaneceu desde então pouco alterado. As quatro classes não-crinóides têm estrutura e função semelhantes em seu WVS. Os holothuroides são provavelmente os mais derivados a nível de classe, porque os seus tentáculos de alimentação bucal são estendidos a partir do canal do anel circunoral e não contêm elementos esqueléticos. O recentemente descoberto Xyloplax (esquerda), um estranho tipo de estrela do mar, tem um arranjo WVS mais único: um novo sistema com canais de anel duplo contendo conexões inter-radiais e um conjunto mono-serial de pés tubulares. Janies e McEdward (1994) sugerem que a geometria única do seu WVS evoluiu através de uma modificação no mecanismo de desenvolvimento de um asteróide juvenil velatid. Este caso representa a evolução mais moderna e derivada do WVS.
A rápida diversificação do equinoderm, a falta de boa preservação e a ausência de mais de um espécime do tipo para comparar classes fósseis obscuras apresenta problemas quando se tenta explicar a sua história evolutiva. A WVS não é, a rigor, apenas uma característica única do equinoderm. Há semelhanças com estruturas nos hemicordados e no fósforo operado hidraulicamente. Sem mencionar que as características embriológicas dos equinodermos também são compartilhadas entre os chordates, hemichordates e os lophophorates. Além disso, o canal do anel desenvolvido a partir de um precursor em forma de U que eventualmente se funde durante a ontogenia para formar o anel que vemos nos equinodermos hoje. Isto parece encaixar com a teoria de Nichols (1967, 1972) que os equinodermos compartilharam uma ancestralidade com um predecessor lophophorate-like, que também têm tripas em forma de U em ambas as formas larval e adulto.
O WVS é uma inovação única e, nos equinodermos de hoje, bem sucedida. Parece plausível que a intrincada rede de canais poderia ter sido construída sobre estruturas anatómicas pré-existentes, como o lophophore e o intestino em forma de U de um antepassado comum em forma de lophophate, mas a evidência não é clara. O que me impressiona é que tal inovação não está presente em outras phyla. A WVS provavelmente surgiu como uma estrutura de alimentação em equinodermos sésseis precoces, como evidenciado por interpretações fósseis, e secundariamente tornou-se usada para locomoção. Outra interpretação pode ser a de que a SVW evoluiu mais de uma vez entre equinodermos extintos e extintos para diferentes fins (i.e. respiração, alimentação, locomoção). De qualquer forma, a WVS é uma adaptação surpreendente entre os equinodermas.

De Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) Uma nova classe de equinodermas da Nova Zelândia. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) The water vascular system and functional morphology of Paleozoic asteroids. Lethaia 21: 189-206.
3) Binyon J. (1972) Physiology of Echinoderms. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) Uma reavaliação da perda de fluido resultante do funcionamento do sistema vascular aquático do asteróide, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrados. Sinauer Associates, Inc. (2003) Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: Vol. IV Echinodermata. McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development. Biol. Touro. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) Uma hipótese para a evolução do sistema Concentriciclóide água-vascular. Em Wilson Jr. W.H., Stricker S.A., Shinn G.L. (eds.): Reprodução e Desenvolvimento dos Invertebrados Marinhos. John Hopkins University Press, Baltimore.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-some implications of the sister-group relationship between echinoderms and chordates. Em Paul C.R.C., Smith A.B. (eds.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, Londres: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) The early phylogeny of chordates and echinoderms and the origin of chordate left-right symmetry and bilateral symmetry. Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms. Croom Helm Ltd., Londres.
13) Lowe C.J., Wray G.A. (1997) Radical alterations in the roles of homeobox genes during echinoderm evolution. Natureza 389: 718-721.
14) Menge B.A. (1982) Effects of feeding on the environment: Asteroidea. Em Jangoux M., Lawrence J.M. (eds.): Echinoderm Nutrition. A.A. Balkema, Roterdão: 521-552.
15) Nichols D. (1967): A origem dos equinodermes. Em Millott N. (ed.): Biologia Echinoderm. Academic Press Inc., London: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontologia 15(4): 519-538.
17) Paul C.R.C. (1977) Evolução dos equinodermos primitivos. Em Hallam A. (ed.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Comentários sobre o sistema vascular aquático, os sulcos alimentares e a ascendência dos equinóides Clypeasteroid. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) Manutenção do volume de fluido no sistema vascular aquático da estrela-do-mar. Nature 262: 577-578.
21) Smith A.B. (1988) Para agrupar ou não agrupar: A posição taxonómica do Xyloplax. Em Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (eds.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Alguns aspectos da evolução em Echinodermata. In de Beer O.R. (ed.): Evolução, Ensaios e Aspectos da Biologia Evolutiva Apresentados a E.S. Goodrich. Oxford University Press, Londres: 287-303.
23) Strathmann R.R. (1975) Limitações sobre a diversidade de formas: Ramificação de sistemas ambulacrais de equinodermos. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: Funções e adaptações moleculares. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Functional bauplan of the echinoderms. Em Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Roterdão: 39-47.
28) Wray G.A. (1999) Genes envolvidos na evolução da morfologia do equinodermes. Em Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 232.
Partilhar o post “Veias de Água”: The Evolution of the Echinoderm Water-Vascular System”
- Shares
 Kevin Zelnio (870 Posts)
Kevin Zelnio (870 Posts)