
Grafiken används med tillstånd. Daniel D. Brown, LaughingMantis.com.
Echinodermer är en av de mest högt avledda djurgrupperna med många arter som viktiga komponenter i flera marina samhällen. De klassificeras genom tre grundläggande gemensamma egenskaper: 1) pentaradial symmetri, 2) skelett bestående av tredimensionella kalcitiska element och 3) förekomst av ett vattenkärlsystem med en yttre öppning. Särskilt detta vattenkärlsystem (WVS) förblir en unik tillämpning av ett hydrauliskt system, som genererar tryckgradienter som kan beordra hundratals små, bedårande rörfötter att krypa över ojämna ytor och livnära sig på sega byten med skal som musslor.
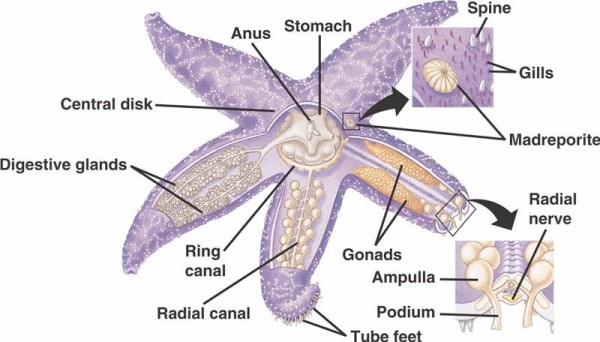
Den grundläggande beståndsdelen i WVS hos icke-krinoida tagghudingar (nedan) omfattar en circumoral ringkanal med fem radiella kanaler som sträcker sig från den. Till ringkanalen hör också madreporiten, som fungerar som en dörr mellan havets yttre miljö och tagghudingens inre miljö. Madreporiten, som fått sitt namn efter sin likhet med korallen Madreporaria (som betyder ”porernas moder”), är fodrad med celler som slår svansliknande flageller i olika riktningar. vätskan inuti WVS liknar havsvatten till sammansättningen men innehåller upp till 60 % mer kaliumjoner, något högre koncentrationer av kloridjoner och fritt svävande albuminhaltigt protein. Man har också funnit att sjögurkor har röda blodkroppar i sin WVS.
Varje radialkanal har seriellt upprepade par av kortare kanaler som fäster vid rörfötterna, vilka drivs av muskelkontraktion av de ampullor som ligger direkt över dem. Tänk dig kalkonbastert: när du trycker på bulben trycker den ut vätska ur bastardens ände, släpp trycket och den suger tillbaka vätska in mot bulben. Crinoiderna skiljer sig åt eftersom de saknar både dessa kortare kanaler och ampullor och deras rörfötter förekommer i grupper om tre som kommer direkt från en kanal som liknar radialkanalen.
Crinoiderna behåller den mest primitiva formen av WVS. Rörfötter tros ursprungligen ha använts helt och hållet för andning samt som matningsstrukturer, men fossila ekorrinoider har tydliga porer mellan skelettplattorna som fick Nichols (1972) att dra slutsatsen att rörfotsystemet inte var särskilt utbrett i ett tidigt skede av crinoidernas evolution. Men han tar inte hänsyn till andra tolkningar av strukturerna, t.ex. öppningar för inre delar som är relaterade till förflyttning eller födosök och som är orelaterade till andning. Karpoider, som Gyrocystis och Dendrocystites, som hittats i medelkambriska sediment anses vara basala för tagghudinglinjen på grund av likheter i skelettelementen och antas ha mjuka, utdragbara inre tentakulära strukturer. Förekomsten av en primitiv WVS är inte klarlagd i fossilregistret av karpoider, men den anses ha utvecklats före andra egenskaper hos tagghudingar. Den tidigaste antydningen av något som liknar en WVS kommer från Helicoplacoidea som hittats i nedre kambriska avlagringar (noterat genom förekomsten av mycket organiserade rader av ambulacralporer). Enligt Paul (1977) har ingen modern klass divergerat från karpoiderna, som har varit utdöda sedan Mesozoikum. Helicoplacoiderna, med triradial symmetri, divergerade mycket tidigt från karpoiderna och resten av stekelhudslinjen utvecklades på Helicoplacoid bauplan.

Xyloplax från Voight 2005.
Sammanställningen av tagghudingar är inte helt utarbetad, men de upplevde en kort diversifieringshändelse under kambrium och ordivicier och den grundläggande bauplanen har sedan dess förblivit föga förändrad. De fyra klasserna som inte är krinoider har liknande struktur och funktion i sina WVS. Holothuroiderna är sannolikt de mest härledda på klassnivå, eftersom deras bucchala födotentakler är förlängda från den circumorala ringkanalen och inte innehåller några skelettelement. Den nyligen upptäckta Xyloplax (till vänster), en märklig sorts sjöstjärna, har ett mer unikt WVS-arrangemang: ett nytt system med dubbla ringkanaler som innehåller interradiella förbindelser och en monoseriell uppsättning rörfötter. Janies och McEdward (1994) föreslår att den unika geometrin hos dess WVS utvecklats via en modifiering i utvecklingsmekanismen hos en juvenil velatid Asteroid. Detta fall representerar den mest moderna och härledda evolutionen av WVS.
Tjurenheternas snabba diversifiering, bristen på bra bevarande och avsaknaden av mer än ett typexemplar för att jämföra obskyra fossilklasser ställer till problem när man försöker förklara deras evolutionära historia. WVS är i strikt mening inte bara en unik egenskap hos tagghudingar. Det finns likheter med strukturer hos hemichordaterna och den hydrauliskt drivna lophophoren. För att inte nämna att embryologiska egenskaper hos tagghudingar också delas av chordaterna, hemichordaterna och lophophoraterna. Dessutom utvecklades ringkanalen från en U-formad föregångare som så småningom smälter samman under ontogenin för att bilda den ring som vi ser hos tagghudingar idag. Detta verkar stämma överens med Nichols’ (1967, 1972) teori om att tagghudingar delade en härstamning med en lophophorate-liknande föregångare, som också har U-formade tarmar i både larv- och vuxenform.
WVS är en unik och, hos dagens tagghudingar, framgångsrik innovation. Det verkar rimligt att det intrikata nätverket av kanaler kan ha byggts på redan existerande anatomiska strukturer, såsom lophophore och U-formad tarm hos en lophophate-liknande gemensam förfader, men bevisen är oklara. Det som är slående för mig är att en sådan innovation inte finns i andra stamceller. WVS uppstod troligen som en födostruktur hos tidiga fastsittande tagghudingar, vilket framgår av fossila tolkningar, och användes i andra hand för att förflytta sig. En annan tolkning kan vara att WVS utvecklades mer än en gång bland utdöda och levande tagghudingar för olika syften (dvs. andning, födosök, förflyttning). Hur som helst är WVS en fantastisk anpassning bland Echinodermata.

Från Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) A new class of Echinodermata from New Zealand. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) The water vascular system and functional morphology of Paleozoic asteroids. Lethaia 21: 189-206.
3) Binyon J. (1972) Physiology of Echinoderms. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) A re-appraisal of the fluid loss resulting from operation of the water vascular system of the starfish, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrates. Sinauer Associates, Inc. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: Vol. IV Echinodermata. McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) En hypotes om utvecklingen av det vattenkärlsmässiga systemet hos Concentricycloid. I Wilson Jr. W.H., Stricker S.A., Shinn G.L. (red.): Reproduction and Development of Marine Invertebrates. John Hopkins University Press, Baltimore.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-some implications of the sister-group relationship between echinoderms and chordates. I Paul C.R.C., Smith A.B. (red.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, London: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) The early phylogeny of chordates and echinoderms and the origin of chordate left-right symmetry and bilateral symmetry. Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms. Croom Helm Ltd., London.
13) Lowe C.J., Wray G.A. (1997) Radikala förändringar i homeobox-genernas roller under utvecklingen av tagghudingar. Nature 389: 718-721.
14) Menge B.A. (1982) Effekter av födan på miljön: Asteroidea. I Jangoux M., Lawrence J.M. (red.): Echinoderm Nutrition. A.A. Balkema, Rotterdam: 521-552.
15) Nichols D. (1967) The origin of echinoderms. I Millott N. (red.): Echinoderm Biology. Academic Press Inc., London: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontology 15(4): 519-538.
17) Paul C.R.C. (1977) Evolution av primitiva tagghudingar. I Hallam A. (red.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Comments on the water vascular system, food grooves, and ancestry of Clypeasteroid Echinoids. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) Upprätthållande av vätskevolym i sjöstjärnans vattenkärlsystem. Nature 262: 577-578.
21) Smith A.B. (1988) To group or not to group: Den taxonomiska positionen för Xyloplax. I Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (red.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Some aspects of evolution in Echinodermata. I de Beer O.R. (red.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, London: 287-303.
23) Strathmann R.R. (1975) Limitations on diversity of forms: Branching of ambulacral systems of echinoderms. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: Funktioner och molekylära anpassningar. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Functional bauplan of the echinoderms. I Candia Carnevali M.D., Bonasoro F. (red.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) Genes involved in the evolution of echinoderm morphology. I Candia Carnevali M.D., Bonasoro F. (red.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 232.
Dela inlägget ”Veins of Water: The Evolution of the Echinoderm Water-Vascular System”
- Shares
 Kevin Zelnio (870 inlägg)
Kevin Zelnio (870 inlägg)