
Grafie použita se svolením. Daniel D. Brown, LaughingMantis.com.
Echinodermy jsou jednou z nejvíce odvozených skupin živočichů s mnoha druhy jako významnými součástmi několika mořských společenstev. Jsou klasifikováni podle tří základních společných znaků: 1) pentaradiální symetrie, 2) kostra z trojrozměrných kalcitických prvků a 3) přítomnost vodního cévního systému s vnějším otvorem. Zejména tento vodní cévní systém (WVS) zůstává jedinečnou aplikací hydraulického systému, vytvářejícího tlakové gradienty, které jsou schopny poručit stovkám drobných, rozkošatělých trubicových nohou plazit se po nerovném povrchu a živit se houževnatou kořistí se skořápkami, jako jsou například mlži.
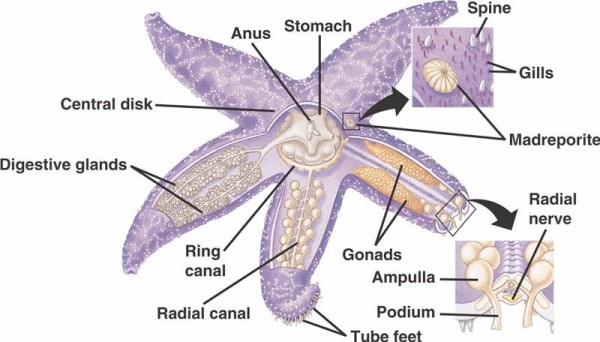
Základní součásti WVS u nekrinoidních ostnokožců (níže) zahrnují cirkumorální prstencový kanál, z něhož vybíhá pět radiálních kanálků. K prstencovému kanálu je rovněž připojen madreporit, který funguje jako brána mezi vnějším prostředím moře a vnitřním prostředím ostnokožce. Madreporit, pojmenovaný podle podobnosti s korálem Madreporaria (což znamená „matka pórů“), je vystlán buňkami, které různými směry bijí ocasními bičíky. tekutina uvnitř WVS je svým složením podobná mořské vodě, ale obsahuje až o 60 % více draselných iontů, o něco vyšší koncentraci chloridových iontů a volně plovoucí bílkovinu albumin. Bylo zjištěno, že mořské okurky mají ve WVS také červené krvinky.
Každý radiální kanálek má sériově se opakující páry kratších kanálků, které se připojují k trubicovým nožkám, jež jsou ovládány svalovou kontrakcí ampulí umístěných přímo nad nimi. Představte si krůtí baňku: když stisknete baňku, vytlačí tekutinu ven z konce baňky, uvolníte tlak a ta nasaje tekutinu zpět směrem k baňce. Krinoidi se liší tím, že postrádají jak tyto kratší kanálky, tak ampuly a jejich trubicové nohy se vyskytují ve skupinách po třech vycházejících přímo z kanálu podobného radiálnímu kanálu.
Krinoidi si zachovali nejprimitivnější formu WVS. Předpokládalo se, že trubicové nohy původně sloužily výhradně k dýchání a také jako potravní struktury, ale fosilní eokrinoidi mají výrazné póry mezi kosterními deskami, což vedlo Nicholse (1972) k závěru, že systém trubicových nohou nebyl na počátku evoluce crinoidů příliš rozsáhlý. Neuvažuje však o jiných interpretacích těchto struktur, jako jsou otvory pro vnitřní části související s pohybem nebo krmením a nesouvisející s dýcháním. Karpoidi, jako Gyrocystis a Dendrocystites, nalezení v sedimentech středního kambria, jsou považováni za bazální k linii ostnokožců kvůli podobnosti kosterních prvků a předpokládá se, že mají měkké, výsuvné vnitřní chapadlové struktury. Existence primitivní WVS není ve fosilním záznamu karpoidů průkazná, přesto se má za to, že se vyvinula dříve než ostatní znaky ostnokožců. Nejstarší náznak něčeho jako WVS pochází od Helicoplacoidea nalezeného ve spodnokambrických usazeninách (vyznačuje se přítomností vysoce organizovaných řad ambulakrálních pórů). Podle Paula (1977) se žádná moderní třída neodlišuje od karpoidů, kteří vymřeli již v druhohorách. Helicoplacoidy s triradiální symetrií se od karpoidů oddělily velmi brzy a zbytek linie ostnokožců se vyvinul na základě helicoplacoidního bauplanu.

Xyloplax z Voight 2005.
Přestože rodokmen ostnokožců není zcela zpracován, prodělali během kambria a ordoviku krátkou diverzifikační událost a základní bauplan se od té doby příliš nezměnil. Čtyři třídy nekrinoidů mají ve svém WVS podobnou strukturu a funkci. Holothuroidi jsou na úrovni třídy pravděpodobně nejodvozenější, protože jejich bukální potravní chapadla jsou prodloužena z cirkumorálního prstencového kanálu a neobsahují žádné kosterní prvky. Nedávno objevený Xyloplax (vlevo), zvláštní druh mořské hvězdice, má unikátnější uspořádání WVS: nový systém s dvojitými prstencovými kanálky obsahujícími interradiální spojení a monoseriální sadu trubicových nožek. Janies a McEdward (1994) se domnívají, že unikátní geometrie jejího WVS se vyvinula díky modifikaci vývojového mechanismu juvenilní velatidy Asteroid. Tento případ představuje nejmodernější a nejodvozenější evoluci WVS.
Rychlá diverzifikace ostnokožců, nedostatek dobrého dochování a absence více než jednoho typového exempláře pro srovnání nejasných fosilních tříd představuje problémy při pokusech o vysvětlení jejich evoluční historie. WVS není, přísně vzato, pouze jedinečným znakem ostnokožců. Existují podobnosti se strukturami u hemichordátů a hydraulicky ovládaných lopoforů. Nemluvě o tom, že embryologické charakteristiky ostnokožců jsou společné i chordátům, hemichordátům a lophoforům. Navíc se prstencovitý kanál vyvinul z předchůdce ve tvaru písmene U, který se nakonec během ontogeneze spojil a vytvořil prstenec, který dnes vidíme u ostnokožců. Zdá se, že to odpovídá Nicholsově (1967, 1972) teorii, že ostnokožci sdíleli předky s předchůdci podobnými lophoforátům, kteří mají také střeva ve tvaru písmene U, a to jak u larválních, tak u dospělých forem.
WVS je jedinečná a u dnešních ostnokožců úspěšná inovace. Zdá se pravděpodobné, že složitá síť kanálů mohla být vybudována na základě již existujících anatomických struktur, jako je lophofor a střevo ve tvaru písmene U u společného předka podobného lophofatu, ale důkazy jsou nejasné. Zarážející pro mě je, že taková inovace není přítomna u jiných fylů. WVS pravděpodobně vznikl jako potravní struktura u raných přisedlých ostnokožců, jak dokládají fosilní interpretace, a druhotně začal sloužit k lokomoci. Jiná interpretace může spočívat v tom, že WVS se u vymřelých a žijících ostnokožců vyvinula vícekrát pro různé účely (tj. dýchání, krmení, lokomoce). Ať tak či onak, WVS je mezi ostnokožci úžasnou adaptací.

Z Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) A new class of Echinodermata from New Zealand. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) The water vascular system and functional morphology of Paleozoic asteroids. Lethaia 21: 189-206.
3) Binyon J. (1972) Physiology of Echinoderms. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) A re-appraisal of the fluid loss resulting from operation of the water vascular system of the starfish, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrates. Sinauer Associates, Inc. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: IV Echinodermata (ostnokožci). McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) A hypothesis for the evolution of the Concentricycloid water-vascular system. In Wilson Jr W.H., Stricker S.A., Shinn G.L. (eds.): Reproduction and Development of Marine Invertebrates. John Hopkins University Press, Baltimore.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-some implications of the sister-group relationship between echinoderms and chordates. In Paul C.R.C., Smith A.B. (eds.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, London: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) The early phylogeny of chordates and echinoderms and the origin of chordate left-right symmetry and bilateral symmetry. Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms (Funkční biologie ostnokožců). Croom Helm Ltd., London.
13) Lowe C.J., Wray G.A. (1997) Radical alterations in the roles of homeobox genes during echinoderm evolution. Nature 389: 718-721.
14) Menge B.A. (1982) Effects of feeding on the environment (Vliv krmení na životní prostředí): Asteroidea. In Jangoux M., Lawrence J.M. (eds.): Echinoderm Nutrition. A.A. Balkema, Rotterdam: 521-552.
15) Nichols D. (1967) The origin of echinoderms. In Millott N. (ed.): Echinoderm Biology. Academic Press Inc, London: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontology 15(4): 519-538.
17) Paul C.R.C. (1977) Evoluce primitivních ostnokožců. In Hallam A. (ed.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Comments on the water vascular system, food grooves, and ancestry of Clypeasteroid Echinoids [Komentáře k vodnímu cévnímu systému, potravním rýhám a původu echinoidů rodu Clypeasteroid]. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) Maintenance of fluid volume in the starfish water vascular system. Nature 262: 577-578.
21) Smith A.B. (1988) To group or not to group: The taxonomic position of Xyloplax. In Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (eds.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Some aspects of evolution in Echinodermata. In de Beer O.R. (ed.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, London: 287-303.
23) Strathmann R.R. (1975) Limitations on diversity of forms: Větvení ambulakrálních systémů ostnokožců. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: Funkce a molekulární adaptace. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Funkční bauplan ostnokožců. In Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) Genes involved in the evolution of echinoderm morphology. In Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 232.
Sdílet příspěvek „Veins of Water: The Evolution of Echinoderm Water-Vascular System“
- Shares
 Kevin Zelnio (870 příspěvků)
Kevin Zelnio (870 příspěvků)