
A grafikát engedéllyel használtuk. Daniel D. Brown, LaughingMantis.com.
A tüskésbőrűek az egyik legmagasabban származtatott állatcsoport, számos fajuk számos tengeri közösség jelentős alkotóeleme. Három alapvető közös jellemzőjük alapján osztályozzák őket: 1) pentaradiális szimmetria, 2) háromdimenziós meszes elemekből álló csontváz, és 3) külső nyílással rendelkező vízi érrendszer jelenléte. Különösen ez a víz-érrendszer (WVS) marad a hidraulikus rendszer egyedülálló alkalmazása, amely olyan nyomásgradienseket hoz létre, amelyek több száz apró, imádnivaló csőlábat képesek parancsolni az egyenetlen felületeken való kúszáshoz és a kemény héjú zsákmányállatokkal, például kagylókkal való táplálkozáshoz.
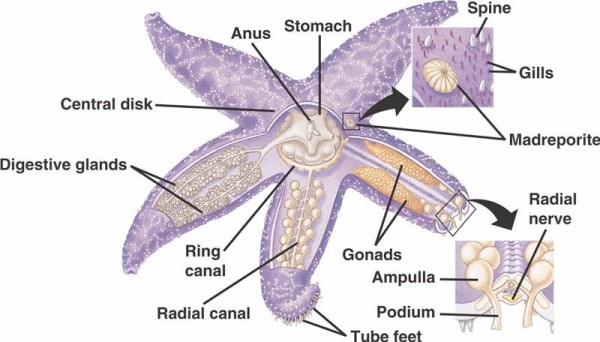
A nem krinoida tüskésbőrűek WVS-ének alapvető összetevői (alább) egy circumoralis gyűrűcsatorna, amelyből öt radiális csatorna nyúlik ki. A gyűrűscsatornához kapcsolódik a madreporita is, amely ajtóként működik a tengeri külső környezet és a tüskésbőrűek belső környezete között. A madreporit, amely a Madreporaria korallhoz való hasonlóságáról kapta a nevét (jelentése “a pórusok anyja”), olyan sejtekkel van bélelve, amelyek különböző irányokba verik a farokszerű flagellákat. a WVS belsejében lévő folyadék összetételében a tengervízhez hasonló, de akár 60%-kal több káliumiont, valamivel magasabb koncentrációban kloridionokat és szabadon lebegő fehérjéket tartalmaz. A tengeri uborkák WVS-ében vörösvérsejteket is találtak.
Minden sugárcsatornának sorban ismétlődő, rövidebb csatornák párjai vannak, amelyek a csőlábakhoz kapcsolódnak, amelyeket a közvetlenül felettük elhelyezkedő ampullák izomösszehúzódása működtet. Gondoljunk a pulykacsőre: ha összenyomjuk a gumót, az folyadékot nyom ki a gumóvégen, ha elengedjük a nyomást, a folyadékot visszaszívja a gumó felé. A tüskésbőrűek abban különböznek, hogy ezekből a rövidebb csatornákból és az ampullákból egyaránt hiányoznak, és a csőlábaik közvetlenül a sugárcsatornához hasonló csatornából kiinduló hármas csoportokban fordulnak elő.
A krinoidák őrzik a WVS legprimitívebb formáját. Úgy vélték, hogy a csőlábakat eredetileg teljes egészében légzésre, valamint táplálkozási szerkezetként használták, de a fosszilis eokrinoidáknál a csontvázlemezek között határozott pórusok vannak, ami Nichols (1972) arra a következtetésre jutott, hogy a csőlábrendszer nem volt túl kiterjedt a crinoidák evolúciójának korai szakaszában. Ő azonban nem veszi figyelembe a szerkezetek egyéb értelmezéseit, például a mozgáshoz vagy táplálkozáshoz kapcsolódó, a légzéshez nem kapcsolódó belső részek nyílásait. A középső kambriumi üledékekben talált karpoidákat, mint például a Gyrocystis és a Dendrocystites, a csontvázelemek hasonlósága miatt a tüskésbőrűek vonalának bazális egyedeinek tekintik, és feltételezik, hogy puha, kihúzható belső csápos szerkezetekkel rendelkeznek. A primitív WVS létezése nem meggyőző a pontyalakúak fosszilis feljegyzéseiben, mégis úgy vélik, hogy a többi tüskésbőrű jellemzőjét megelőzően fejlődött ki. A WVS-re utaló legkorábbi utalást az alsó kambriumi lerakódásokban talált Helicoplacoidea (az ambulakrális pórusok magasan szervezett sorainak jelenlétéről ismert). Paul (1977) szerint egyetlen modern osztály sem vált el a carpoidáktól, amelyek a mezozoikum óta kihaltak. A triradiális szimmetriájú helicoplacoidák nagyon korán elváltak a carpoidáktól, és a többi tüskésbőrű vonal a helicoplacoidák bauplanján fejlődött ki.

Xyloplax a Voight 2005-ből.
Míg a tüskésbőrűek családfája nem teljesen kidolgozott, a kambrium és az ordivícium során rövid diverzifikációs eseményen mentek keresztül, és az alap bauplan azóta is alig változott. A négy nem krinoid osztály WVS-ének szerkezete és funkciója hasonló. A holothuroidák valószínűleg a legjobban származtatottak az osztály szintjén, mivel a bukkális táplálkozó csápjaik a circumoralis gyűrűcsatornából nyúlnak ki, és nem tartalmaznak csontvázelemeket. A nemrégiben felfedezett Xyloplax (balra), egy különös tengeri csillagfajta, sokkal egyedibb WVS-elrendezéssel rendelkezik: újszerű rendszer kettős gyűrűscsatornákkal, amelyek interradialis kapcsolatokat tartalmaznak, és egyszériás csőlábakkal rendelkeznek. Janies és McEdward (1994) szerint a WVS-ének egyedi geometriája egy juvenilis velatid Asteroid fejlődési mechanizmusának módosítása révén alakult ki. Ez az eset képviseli a WVS legmodernebb és származtatott evolúcióját.
A tüskésbőrűek gyors diverzifikációja, a jó megőrzés hiánya és az egynél több típuspéldány hiánya a homályos fosszilis osztályok összehasonlításához problémát jelent, amikor megpróbáljuk megmagyarázni evolúciós történetüket. A WVS szigorúan véve nem csak a tüskésbőrűek egyedi jellemzője. Vannak hasonlóságok a hemichordaták és a hidraulikusan működő lophore struktúráival. Arról nem is beszélve, hogy a tüskésbőrűek embriológiai jellemzői közösek a gerincesek, a hemichordaták és a lophophoraták között is. Ráadásul a gyűrűcsatorna egy U alakú előfutárból fejlődött ki, amely az ontogenezis során végül összeolvad, és kialakítja a tüskésbőrűeknél ma látható gyűrűt. Ez illeszkedni látszik Nichols (1967, 1972) elméletéhez, miszerint a tüskésbőrűek közös ősökkel rendelkeztek egy lophophorata-szerű előddel, amelyeknek szintén U-alakú belük van mind a lárva-, mind a kifejlett formában.
A WVS egyedülálló és a mai tüskésbőrűeknél sikeres innováció. Valószínűnek tűnik, hogy a csatornák bonyolult hálózata már létező anatómiai struktúrákra épülhetett, mint például a lophophophore és az U alakú bél egy lophophore-szerű közös ősre, de a bizonyítékok nem egyértelműek. Ami számomra szembetűnő, az az, hogy egy ilyen újítás más törzseknél nincs jelen. A WVS valószínűleg táplálkozási struktúraként alakult ki a korai ülő tüskésbőrűeknél, amint azt a fosszilis értelmezések bizonyítják, és másodlagosan a helyváltoztatásra használták. Egy másik értelmezés szerint a WVS többször is kialakult a kihalt és a ma élő tüskésbőrűek között különböző célokra (azaz légzésre, táplálkozásra, mozgásra). Akárhogy is, a WVS egy elképesztő alkalmazkodás az Echinodermata között.

From Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) A new class of Echinodermata from New Zealand. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) The water vascular system and functional morphology of Paleozoic asteroids. Lethaia 21: 189-206.
3) Binyon J. (1972) Physiology of Echinoderms. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) A reappraisal of the fluid loss resulting from operation of the water vascular system of the starfish, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrates. Sinauer Associates, Inc. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: Vol. IV Echinodermata. McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) A Concentricycloid víz-vaszkuláris rendszer evolúciójának hipotézise. In Wilson Jr. W.H., Stricker S.A., Shinn G.L. (szerk.): Reproduction and Development of Marine Invertebrates. John Hopkins University Press, Baltimore.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-some implications of the sister-group relationship between echinoderms and chordates. In Paul C.R.C., Smith A.B. (szerk.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, London: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) The early phylogeny of chordates and echinoderms and the origin of chordate left-right symmetry and bilateral symmetry. Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms. Croom Helm Ltd., London.
13) Lowe C.J., Wray G.A. (1997) Radical alterations in the roles of homeobox genes during echinoderm evolution. Nature 389: 718-721.
14) Menge B.A. (1982) A táplálkozás hatása a környezetre: Asteroidea. In Jangoux M., Lawrence J.M. (szerk.): Echinoderm Nutrition. A.A. Balkema, Rotterdam: 521-552.
15) Nichols D. (1967) The origin of echinoderms. In Millott N. (szerk.): Echinoderm Biology. Academic Press Inc., London: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontology 15(4): 519-538.
17) Paul C.R.C. (1977) A primitív tüskésbőrűek evolúciója. In Hallam A. (szerk.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Megjegyzések a Clypeasteroid Echinoidák vízi érrendszeréről, táplálékbarázdáiról és ősiségéről. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) A folyadékmennyiség fenntartása a tengeri csillagok vízi érrendszerében. Nature 262: 577-578.
21) Smith A.B. (1988) To group or not to group: A Xyloplax rendszertani helyzete. In Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (szerk.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Some aspects of evolution in Echinodermata. In de Beer O.R. (szerk.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, London: 287-303.
23) Strathmann R.R. (1975) Limitations on diversity of forms: A tüskésbőrűek ambulakrális rendszereinek elágazása. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: Funkciók és molekuláris adaptációk. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Functional bauplan of the echinoderms. In Candia Carnevali M.D., Bonasoro F. (szerk.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) Genes involved in the evolution of echinoderm morphology. In Candia Carnevali M.D., Bonasoro F. (szerk.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 232.
Share the post “Veins of Water: The Evolution of the Echinoderm Water-Vascular System”
- Shares
 Kevin Zelnio (870 hozzászólás)
Kevin Zelnio (870 hozzászólás)