
Gráfico utilizado con permiso. Daniel D. Brown, LaughingMantis.com.
Los equinodermos son uno de los grupos de animales más derivados, con muchas especies como componentes significativos de varias comunidades marinas. Se clasifican por tres características fundamentales compartidas: 1) simetría pentaradial, 2) esqueleto formado por elementos calcíticos tridimensionales, y 3) la presencia de un sistema vascular acuático con una abertura externa. Este sistema vascular acuático (WVS), en particular, sigue siendo una aplicación única de un sistema hidráulico, generando gradientes de presión que son capaces de comandar cientos de diminutos y adorables pies tubulares para arrastrarse sobre superficies irregulares y alimentarse de presas duras con concha como los mejillones.
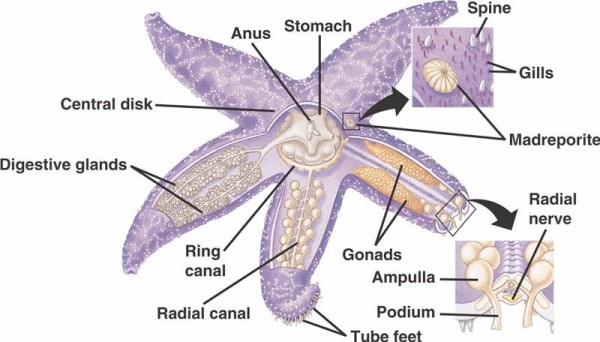
Los componentes básicos del WVS en los equinodermos no crinoideos (abajo) incluyen un canal anular circunoral con cinco canales radiales que se extienden desde él. Además, el canal anular está unido a la madreporita, que actúa como una puerta entre el medio externo del mar y el medio interno del equinodermo. El madreporito, llamado así por su parecido con el coral Madreporaria (que significa «madre de los poros»), está revestido de células que baten flagelos en forma de cola en diversas direcciones.El fluido del interior del VSM es similar al agua de mar en su composición, pero contiene hasta un 60% más de iones de potasio, concentraciones ligeramente superiores de iones de cloruro y proteínas albuminosas que flotan libremente. También se ha encontrado que los pepinos de mar tienen glóbulos rojos en su VSM.
Cada canal radial tiene pares de canales más cortos que se repiten en serie y que se unen a los pies tubulares, los cuales son operados por la contracción muscular de las ampollas ubicadas directamente sobre ellos. Piensa en una ampolla de pavo: cuando aprietas la ampolla empuja el líquido hacia el extremo de la ampolla, deja de presionar y vuelve a succionar el líquido hacia la ampolla. Los crinoideos se diferencian porque carecen tanto de estos canales más cortos como de las ampollas y sus pies tubulares se presentan en grupos de tres saliendo directamente de un canal similar al canal radial.
Los crinoideos conservan la forma más primitiva del VSM. Se creía que los pies tubulares se utilizaban originalmente en su totalidad para la respiración, así como las estructuras de alimentación, pero los eocrinoideos fósiles tienen poros distintivos entre las placas del esqueleto que llevaron a Nichols (1972) a concluir que el sistema de pies tubulares no era muy extenso en las primeras etapas de la evolución de los crinoideos. Pero no tiene en cuenta otras interpretaciones de las estructuras, como aberturas para partes internas relacionadas con la locomoción o la alimentación y no relacionadas con la respiración. Los carpoides, como Gyrocystis y Dendrocystites, encontrados en sedimentos del Cámbrico medio, se consideran basales al linaje de los equinodermos debido a la similitud en los elementos del esqueleto y se hipotetiza que tienen estructuras tentaculares internas blandas y protráctiles. La existencia de un VSM primitivo no es concluyente en el registro fósil de los carpoides, pero se considera que evolucionó antes que otras características de los equinodermos. La primera sugerencia de algo parecido a un SVA proviene de los Helicoplacoidea encontrados en los depósitos del Cámbrico inferior (que se caracterizan por la presencia de hileras de poros ambulacrales muy organizados). Según Paul (1977) ninguna clase moderna ha divergido de los carpoideos, extinguidos desde el Mesozoico. Los helicoplacoides, con simetría trirradial, divergieron muy pronto de los carpoides y el resto del linaje de equinodermos evolucionó sobre el bauplano de los helicoplacoides.

Xyloplax de Voight 2005.
Aunque el árbol genealógico de los equinodermos no está del todo resuelto, experimentaron un breve evento de diversificación durante el Cámbrico y el Ordiviano y el bauplan básico ha permanecido desde entonces poco modificado. Las cuatro clases de no-crinoideos tienen una estructura y función similares en su VSM. Los holoturoideos son probablemente los más derivados a nivel de clase, porque sus tentáculos de alimentación bucal se extienden desde el canal anular circunoral y no contienen elementos esqueléticos. El recientemente descubierto Xyloplax (izquierda), una extraña especie de estrella de mar, tiene una disposición de WVS más singular: un sistema novedoso con canales anulares dobles que contienen conexiones interradiales y un conjunto monoserial de pies tubulares. Janies y McEdward (1994) sugieren que la geometría única de su WVS evolucionó a través de una modificación en el mecanismo de desarrollo de un asteroide velatidio juvenil. Este caso representa la evolución más moderna y derivada del VSM.
La rápida diversificación de los equinodermos, la falta de una buena conservación y la ausencia de más de un espécimen tipo para comparar clases fósiles oscuras presenta problemas a la hora de intentar explicar su historia evolutiva. El WVS no es, estrictamente hablando, sólo una característica única de los equinodermos. Hay similitudes con las estructuras de los hemicordados y el lofóforo de funcionamiento hidráulico. No hay que olvidar que las características embriológicas de los equinodermos también se comparten entre los cordados, los hemicordados y los loforados. Además, el canal anular se desarrolló a partir de un precursor en forma de U que finalmente se fusiona durante la ontogenia para formar el anillo que vemos en los equinodermos actualmente. Esto parece encajar con la teoría de Nichols (1967, 1972) de que los equinodermos compartieron una ascendencia con un predecesor similar a los lofoforados, que también tienen intestinos en forma de U tanto en las formas larvarias como en las adultas.
El WVS es una innovación única y, en los equinodermos actuales, exitosa. Parece plausible que la intrincada red de canales pudiera haberse construido sobre estructuras anatómicas preexistentes, como el lofóforo y el intestino en forma de U de un ancestro común lofato, pero las pruebas no son claras. Lo que me llama la atención es que tal innovación no está presente en otros filos. El VSM probablemente surgió como una estructura de alimentación en los primeros equinodermos sésiles, como demuestran las interpretaciones de los fósiles, y en segundo lugar se utilizó para la locomoción. Otra interpretación puede ser que el VSM evolucionó más de una vez entre los equinodermos extintos y existentes para diferentes propósitos (es decir, respiración, alimentación, locomoción). Cualquier manera, el WVS es una adaptación asombrosa entre el Echinodermata.

De Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) Una nueva clase de Echinodermata de Nueva Zelanda. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) The water vascular system and functional morphology of Paleozoic asteroids. Lethaia 21: 189-206.
3) Binyon J. (1972) Physiology of Echinoderms. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) A re-appraisal of the fluid loss resulting from operation of the water vascular system of the starfish, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrados. Sinauer Associates, Inc. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: Vol. IV Echinodermata. McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) Una hipótesis para la evolución del sistema acuático-vascular concéntrico. En Wilson Jr. W.H., Stricker S.A., Shinn G.L. (eds.): Reproduction and Development of Marine Invertebrates. John Hopkins University Press, Baltimore.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-some implications of the sister-group relationship between echinoderms and chordates. En Paul C.R.C., Smith A.B. (eds.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, Londres: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) The early phylogeny of chordates and echinoderms and the origin of chordate left-right symmetry and bilateral symmetry. Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms. Croom Helm Ltd., Londres.
13) Lowe C.J., Wray G.A. (1997) Radical alterations in the roles of homeobox genes during echinoderm evolution. Nature 389: 718-721.
14) Menge B.A. (1982) Efectos de la alimentación en el medio ambiente: Asteroidea. En Jangoux M., Lawrence J.M. (eds.): Echinoderm Nutrition. A.A. Balkema, Rotterdam: 521-552.
15) Nichols D. (1967) The origin of echinoderms. En Millott N. (ed.): Echinoderm Biology. Academic Press Inc., Londres: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontology 15(4): 519-538.
17) Paul C.R.C. (1977) Evolution of primitive echinoderms. En Hallam A. (ed.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Comments on the water vascular system, food grooves, and ancestry of Clypeasteroid Echinoids. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) Maintenance of fluid volume in the starfish water vascular system. Nature 262: 577-578.
21) Smith A.B. (1988) To group or not to group: La posición taxonómica de Xyloplax. En Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (eds.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Some aspects of evolution in Echinodermata. En de Beer O.R. (ed.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, Londres: 287-303.
23) Strathmann R.R. (1975) Limitations on diversity of forms: La ramificación de los sistemas ambulacrales de los equinodermos. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: Funciones y adaptaciones moleculares. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Functional bauplan of the echinoderms. En Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) Genes implicados en la evolución de la morfología de equinodermo. En Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 232.
Comparte el post «Venas de agua: La evolución del sistema hidrovascular de los equinodermos»
- Shares
 Kevin Zelnio (870 Mensajes)
Kevin Zelnio (870 Mensajes)