
Grafic folosit cu permisiune. Daniel D. Brown, LaughingMantis.com.
Echinodermele sunt unul dintre cele mai înalt derivate grupuri de animale, multe specii fiind componente semnificative ale mai multor comunități marine. Ele sunt clasificate după trei caracteristici fundamentale comune: 1) simetria pentaradiană, 2) scheletul alcătuit din elemente calcitice tridimensionale și 3) prezența unui sistem vascular acvatic cu deschidere externă. Acest sistem vascular acvatic (WVS), în special, rămâne o aplicație unică a unui sistem hidraulic, generând gradienți de presiune care sunt capabili să comande sute de picioare tubulare minuscule, adorabile, pentru a se târî pe suprafețe inegale și a se hrăni cu prăzi cu cochilii rezistente, cum ar fi midii.
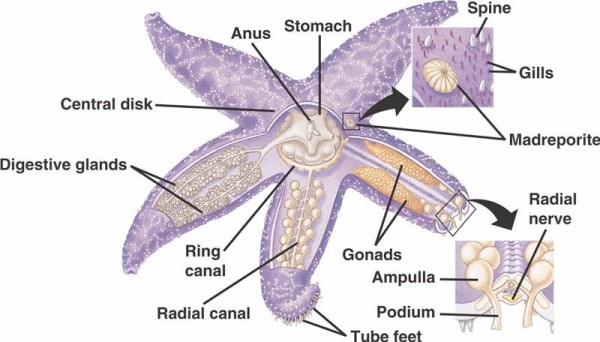
Componentele de bază ale WVS la echinodermele necrinoide (mai jos) includ un canal inelar circumoral cu cinci canale radiale care se extind din afara acestuia. De asemenea, atașat de canalul inelar este madreporitul, care acționează ca o ușă între mediul extern al mării și mediul intern al echinodermului. Madreporitul, denumit astfel după asemănarea sa cu coralul Madreporaria (care înseamnă „mama porilor”), este căptușit cu celule care bat flageli în formă de coadă într-o varietate de direcții.Fluidul din interiorul WVS are o compoziție similară cu cea a apei de mare, dar conține cu până la 60% mai mulți ioni de potasiu, concentrații ușor mai mari de ioni de clorură și proteine albuminoase care plutesc liber. S-a descoperit, de asemenea, că castraveții de mare au globule roșii în WVS.
Care canal radial are perechi de canale mai scurte care se repetă în serie și care se atașează la picioarele tubulare, care sunt acționate prin contracția musculară a ampulelor situate direct deasupra lor. Gândiți-vă la un bețișor de curcan: când strângeți bulbul, acesta împinge lichidul în afara capătului bețișorului, lăsați presiunea și acesta aspiră lichidul înapoi spre bulb. Crinoidele diferă pentru că le lipsesc atât aceste canale mai scurte, cât și ampullele, iar picioarele lor tubulare apar în grupuri de trei, venind direct dintr-un canal asemănător canalului radial.
Crinoidele păstrează cea mai primitivă formă a WVS. Se credea că inițial picioarele tubulare au fost folosite în întregime pentru respirație, precum și ca structuri de hrănire, dar eocrinoidele fosile au pori distincți între plăcile scheletului care l-au determinat pe Nichols (1972) să concluzioneze că sistemul picioarelor tubulare nu a fost foarte extins la începutul evoluției crinoidelor. Dar el nu ia în considerare alte interpretări ale structurilor, cum ar fi deschideri pentru părți interne legate de locomoție sau hrănire și care nu au legătură cu respirația. Carpoidele, cum ar fi Gyrocystis și Dendrocystites, găsite în sedimentele din Cambrianul mijlociu, sunt considerate a fi la baza neamului echinodermelor datorită similitudinii elementelor scheletice și se presupune că au structuri tentaculare interne moi și protractibile. Existența unui WVS primitiv nu este concludentă în înregistrările fosile ale carpoidelor, însă se consideră că acesta a evoluat înaintea altor caracteristici ale echinodermelor. Cea mai veche sugestie de ceva asemănător cu un WVS provine de la Helicoplacoidea găsită în depozitele cambriene inferioare (observată prin prezența unor rânduri foarte bine organizate de pori ambulacrali). Potrivit lui Paul (1977), nicio clasă modernă nu a deviat de la carpoide, care a dispărut încă din Mesozoic. Helicoplacoidele, cu simetrie triradială, au deviat foarte devreme de la carpoide, iar restul liniei de echinoderme a evoluat pe bauplanul Helicoplacoidelor.

Xyloplax din Voight 2005.
În timp ce arborele genealogic al echinodermelor nu este în întregime elaborat, acestea au cunoscut un scurt eveniment de diversificare în timpul Cambrianului și Ordivicianului, iar bauplanul de bază a rămas de atunci puțin schimbat. Cele patru clase de necrinoide au o structură și o funcție similară în WVS. Holothuroidele sunt probabil cele mai derivate la nivelul clasei, deoarece tentaculele lor bucale de hrănire sunt extinse din canalul inelar circumoral și nu conțin elemente scheletice. Recent descoperitul Xyloplax (stânga), un fel ciudat de stea de mare, are un aranjament WVS mai unic: un sistem nou cu canale inelare duble care conțin conexiuni inter-radiale și un set monoserial de picioare tubulare. Janies și McEdward (1994) sugerează că geometria unică a WVS-ului său a evoluat prin intermediul unei modificări în mecanismul de dezvoltare a unui asteroid velatid juvenil. Acest caz reprezintă cea mai modernă și derivată evoluție a WVS.
Diversificarea rapidă a echinodermelor, lipsa unei bune conservări și absența a mai mult de un specimen tip pentru compararea claselor fosile obscure prezintă probleme atunci când se încearcă să se explice istoria evoluției lor. WVS nu este, strict vorbind, doar o caracteristică unică a echinodermelor. Există asemănări cu structurile din hemicordați și cu lophoforul cu funcționare hidraulică. Ca să nu mai vorbim de faptul că caracteristicile embriologice ale echinodermelor sunt, de asemenea, împărtășite între cordate, hemicordate și lioforate. În plus, canalul inelar s-a dezvoltat pornind de la un precursor în formă de U care, în cele din urmă, fuzionează împreună în timpul ontogenezei pentru a forma inelul pe care îl vedem astăzi la echinoderme. Acest lucru pare să se potrivească cu teoria lui Nichols (1967, 1972), conform căreia echinodermele au împărtășit un strămoș cu un predecesor asemănător lioforatelor, care au, de asemenea, intestine în formă de U atât la formele larvare, cât și la cele adulte.
WVS este o inovație unică și, la echinodermele de astăzi, o inovație de succes. Pare plauzibil că rețeaua complexă de canale ar fi putut fi construită pe structuri anatomice preexistente, cum ar fi lofoforul și intestinul în formă de U al unui strămoș comun asemănător liofatului, dar dovezile sunt neclare. Ceea ce mă surprinde este faptul că o astfel de inovație nu este prezentă în alte phyla. Probabil că WVS a apărut ca structură de hrănire la echinodermele sesile timpurii, așa cum reiese din interpretările fosilelor, iar în al doilea rând a devenit folosită pentru locomoție. O altă interpretare ar putea fi aceea că WVS a evoluat de mai multe ori la echinoderme dispărute și existente în scopuri diferite (de exemplu, respirație, hrănire, locomoție). Oricum ar fi, WVS este o adaptare uimitoare printre Echinodermata.

De la Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) O nouă clasă de Echinodermata din Noua Zeelandă. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) Sistemul vascular acvatic și morfologia funcțională a asteroizilor paleozoici. Lethaia 21: 189-206.
3) Binyon J. (1972) Fiziologia echinodermelor. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) O reevaluare a pierderilor de fluide rezultate din funcționarea sistemului vascular acvatic al stelei de mare, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrate. Sinauer Associates, Inc. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: Vol. IV Echinodermata. McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Coelomic Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) A hypothesis for the evolution of the Concentricycloid water-vascular system. În Wilson Jr. W.H., Stricker S.A., Shinn G.L. (eds.): Reproduction and Development of Marine Invertebrates. John Hopkins University Press, Baltimore.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-some implications of the sister-group relationship between echinoderms and chordates. În Paul C.R.C., Paul C.R.C., Smith A.B. (eds.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, Londra: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) The early phylogeny of chordates and echinoderms and the origin of chordate left-right symmetry and bilateral symmetry (Filogenia timpurie a chordatelor și echinodermelor și originea simetriei stânga-dreapta și a simetriei bilaterale). Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms (O biologie funcțională a echinodermelor). Croom Helm Ltd., Londra.
13) Lowe C.J., Wray G.A. (1997) Modificări radicale în rolurile genelor homeobox în timpul evoluției echinodermelor. Nature 389: 718-721.
14) Menge B.A. (1982) Efectele hrănirii asupra mediului: Asteroidea. În Jangoux M., Lawrence J.M. (eds.): Echinoderm Nutrition. A.A. Balkema, Rotterdam: 521-552.
15) Nichols D. (1967) The origin of echinoderms. În Millott N. (ed.): Echinoderm Biology. Academic Press Inc., Londra: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontologie 15(4): 519-538.
17) Paul C.R.C. (1977) Evoluția echinodermelor primitive. În Hallam A. (ed.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Comentarii privind sistemul vascular acvatic, canelurile de hrană și strămoșii echinoidelor Clypeasteroid. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) Menținerea volumului de fluid în sistemul vascular acvatic al stelei de mare. Nature 262: 577-578.
21) Smith A.B. (1988) To group or not to group: The taxonomic position of Xyloplax. În Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (eds.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Some aspects of evolution in Echinodermata. În de Beer O.R. (ed.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, London: 287-303.
23) Strathmann R.R. (1975) Limitations on diversity of forms: Ramificarea sistemelor ambulacrale ale echinodermelor. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: Funcții și adaptări moleculare. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Bauplan funcțional al echinodermelor. În Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) Genele implicate în evoluția morfologiei echinodermelor. În Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 232.
Share the post „Venele de apă: The Evolution of the Echinoderm Water-Vascular System”
- Shares
 Kevin Zelnio (870 Posts)
Kevin Zelnio (870 Posts)