
Kuvaa käytetty luvalla. Daniel D. Brown, LaughingMantis.com.
Piikkinahkaiset ovat yksi korkeimmin johdetuista eläinryhmistä, joiden monet lajit ovat merkittäviä osia useissa meriyhteisöissä. Ne luokitellaan kolmen yhteisen perusominaisuuden perusteella: 1) pentaradiaalinen symmetria, 2) kolmiulotteisista kalkkielementeistä koostuva luuranko ja 3) ulkoisella aukolla varustetun veden verisuoniston olemassaolo. Erityisesti tämä vesivaskulaarinen järjestelmä (WVS) on edelleen ainutlaatuinen hydraulisen järjestelmän sovellus, joka tuottaa painegradientteja, jotka kykenevät komentamaan satoja pieniä, ihailtavia putkijalkoja ryömimään epätasaisilla pinnoilla ja syömään sitkeitä kuorellisia saaliseläimiä, kuten simpukoita.
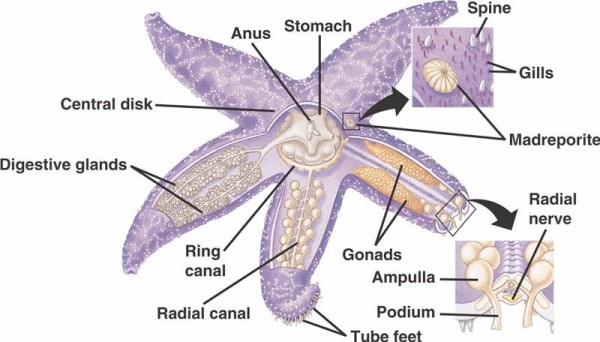
Ei-krinoidien piikkinahkaisten WVS:n peruskomponentteihin (alla) kuuluu circumoraalinen rengasmainen kanava, josta lähtee ulospäin viisi säteittäistä kanavaa. Rengaskanavaan on kiinnittynyt myös madreporiitti, joka toimii ovena meren ulkoisen ympäristön ja piikkinahkaisen sisäisen ympäristön välillä. Madreporiitti, joka on saanut nimensä Madreporaria-korallia (joka tarkoittaa ”huokosten äitiä”) muistuttavan nimensä mukaan, on vuorattu soluilla, jotka lyövät pyrstönmuotoisia lippulankoja eri suuntiin.WVS:n sisällä oleva neste on koostumukseltaan samanlaista kuin merivesi, mutta se sisältää jopa 60 prosenttia enemmän kaliumioneja, hieman suurempia pitoisuuksia kloridi-ioneja ja vapaasti kelluvia albumiinipitoisia proteiineja. Merikurkkujen WVS:ssä on havaittu olevan myös punasoluja.
Jokaiseen säteittäiseen kanavaan kuuluu peräkkäin toistuvia pareja lyhyempiä kanavia, jotka kiinnittyvät putkijalkoihin, joita käytetään suoraan niiden yläpuolella sijaitsevien ampullien lihassupistuksilla. Ajattele kalkkunapuristinta: kun puristat lamppua, se työntää nestettä ulos puristimen päästä, päästä irti paineesta ja se imee nestettä takaisin lamppua kohti. Crinoidit eroavat toisistaan, koska niiltä puuttuvat sekä nämä lyhyemmät kanavat että ampullat, ja niiden putkijalat esiintyvät kolmen putken ryhmissä, jotka tulevat suoraan sädekanavan kaltaisesta kanavasta.
Crinoideilla on säilynyt WVS:n alkeellisin muoto. Putkijalkoja uskottiin alun perin käytetyn kokonaan hengitykseen sekä ravintorakenteina, mutta fossiilisilla eokrinoideilla on luurankolevyjen välissä selvät huokoset, jotka saivat Nicholsin (1972) päättelemään, että putkijalkajärjestelmä ei ollut kovin laaja krinoidien evoluution alkuvaiheessa. Hän ei kuitenkaan ota huomioon muita tulkintoja rakenteista, kuten aukkoja sisäisille osille, jotka liittyvät liikkumiseen tai ruokailuun ja jotka eivät liity hengitykseen. Keskikambrikauden sedimenteistä löytyneiden karpoidien, kuten Gyrocystis ja Dendrocystites, katsotaan luurankoelementtien samankaltaisuuden vuoksi olevan piikkinahkaisten sukulinjan pohjaläheisiä, ja niillä oletetaan olevan pehmeitä, ulosvedettäviä sisäisiä lonkerorakenteita. Alkeellisen WVS:n olemassaolosta ei ole varmuutta karpoidien fossiileissa, mutta sen katsotaan kehittyneen ennen muita piikkinahkaisten ominaisuuksia. Varhaisimmat viitteet WVS:n kaltaisesta rakenteesta ovat peräisin alemman kambrikauden kerrostumista löydetyistä Helicoplacoidea-sukuisista eläimistä (jotka erottuvat hyvin järjestäytyneistä ambulakraalihuokosten riveistä). Paulin (1977) mukaan mikään nykyaikainen luokka ei ole eronnut karpoidista, joka on kuollut sukupuuttoon mesotsooisesta lähtien. Helicoplacoidit, joilla on triradiaalinen symmetria, erosivat hyvin varhain karpoidista, ja muu piikkinahkaisten suku kehittyi helicoplacoidien bauplanin pohjalta.

Xyloplax lähteestä Voight 2005.
Vaikka piikkinahkaisten sukupuu ei ole täysin selvillä, ne kokivat lyhyen diversifikaatiotapahtuman kambrikauden ja ordivikumin aikana, ja sen jälkeen perusbauplani on pysynyt vähän muuttuneena. Neljällä muulla kuin sarvikuonoluokalla on samanlainen rakenne ja toiminta WVS:ssä. Holothuroidit ovat todennäköisesti luokkatasolla kaikkein johdetuimpia, koska niiden bucchal-ruokintalonkerot ulottuvat circumoraalisesta rengaskanavasta, eikä niissä ole luurankoelementtejä. Hiljattain löydetyllä Xyloplaxilla (vasemmalla), oudolla meritähdellä, on ainutlaatuisempi WVS-järjestely: se on uudenlainen järjestelmä, jossa on kaksinkertaiset rengaskanavat, jotka sisältävät radiaalisten yhteyksien välisiä yhteyksiä, ja yksisarjaiset putkijalat. Janies ja McEdward (1994) esittävät, että sen WVS:n ainutlaatuinen geometria on kehittynyt juveniilin velatidi-asteroidin kehitysmekanismin muutoksen kautta. Tämä tapaus edustaa WVS:n nykyaikaisinta ja johdetuinta evoluutiota.
Kotiloiden nopea diversifioituminen, hyvän säilyvyyden puute ja useamman kuin yhden tyyppinäytteen puuttuminen hämäräperäisten fossiililuokkien vertailemiseksi aiheuttavat ongelmia, kun yritetään selittää niiden kehityshistoriaa. WVS ei varsinaisesti ole vain piikkinahkaisten ainutlaatuinen ominaisuus. Siinä on yhtäläisyyksiä hemichordatien rakenteisiin ja hydraulisesti toimiviin lofooreihin. Puhumattakaan siitä, että piikkinahkaisten embryologiset piirteet ovat yhteisiä myös niveljalkaisten, hemichordattien ja lofoorattien kanssa. Lisäksi rengaskanava kehittyi U:n muotoisesta esiasteesta, joka lopulta sulautui yhteen ontogeneesin aikana muodostaen renkaan, jonka näemme piikkinahkaisissa nykyään. Tämä näyttää sopivan yhteen Nicholsin (1967, 1972) teorian kanssa, jonka mukaan piikkinahkaiset jakoivat esi-isänsä lophophoratien kaltaisen edeltäjän kanssa, joilla on myös U:n muotoinen suoli sekä toukka- että aikuismuodossa.
WVS on ainutlaatuinen ja nykyisissä piikkinahkaisissa onnistunut innovaatio. Vaikuttaa uskottavalta, että monimutkainen kanavaverkosto on voinut rakentua jo olemassa olevien anatomisten rakenteiden, kuten lofoforin kaltaisen yhteisen esi-isän lofoforin ja U:n muotoisen suolen, varaan, mutta todisteet ovat epäselviä. Minulle on silmiinpistävää, että tällaista innovaatiota ei esiinny muissa heimoissa. WVS syntyi todennäköisesti ravintorakenteeksi varhaisissa sessiileissä piikkinahkaisissa, kuten fossiilitulkinnat osoittavat, ja sitä käytettiin toissijaisesti liikkumiseen. Toinen tulkinta voi olla, että WVS on kehittynyt useammin kuin kerran sukupuuttoon kuolleiden ja elossa olevien piikkinahkaisten keskuudessa eri tarkoituksiin (eli hengitykseen, ravintoon ja liikkumiseen). Oli miten oli, WVS on hämmästyttävä sopeutuminen piikkinahkaisten keskuudessa.

Vasta Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) A new class of Echinodermata from New Zealand. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) The water vascular system and functional morphology of Paleozoic asteroids. Lethaia 21: 189-206.
3) Binyon J. (1972) Physiology of Echinoderms. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) A re-appraisal of the fluid loss resulting from operation of the water vascular system of the starfish, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrates. Sinauer Associates, Inc. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: Vol. IV Echinodermata. McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) Hypoteesi Concentricycloidin vesivaskulaarisen järjestelmän evoluutiosta. Teoksessa Wilson Jr. W.H., Stricker S.A., Shinn G.L. (toim.): Reproduction and Development of Marine Invertebrates. John Hopkins University Press, Baltimore.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-some implications of the sister-group relationship between echinoderms and chordates. Teoksessa Paul C.R.C., Smith A.B. (toim.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, London: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) The early phylogeny of chordates and echinoderms and the origin of chordate left-right symmetry and bilateral symmetry. Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms. Croom Helm Ltd., Lontoo.
13) Lowe C.J., Wray G.A. (1997) Homeobox-geenien roolien radikaalit muutokset piikkinahkaisten evoluution aikana. Nature 389: 718-721.
14) Menge B.A. (1982) Ruokinnan vaikutukset ympäristöön: Asteroidea. Teoksessa Jangoux M., Lawrence J.M. (toim.): Echinoderm Nutrition. A.A. Balkema, Rotterdam: 521-552.
15) Nichols D. (1967) The origin of echinoderms. Teoksessa Millott N. (toim.): Echinoderm Biology. Academic Press Inc., London: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontology 15(4): 519-538.
17) Paul C.R.C. (1977) Alkukantaisten piikkinahkaisten evoluutio. Teoksessa Hallam A. (toim.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Comments on the water vascular system, food grooves, and ancestry of Clypeasteroid Echinoids. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) Nesteen tilavuuden ylläpito meritähden vesiverisuonistossa. Nature 262: 577-578.
21) Smith A.B. (1988) Ryhmittyä vai olla ryhmittymättä: Xyloplaxin taksonominen asema. Teoksessa Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (toim.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Some aspects of evolution in Echinodermata. Teoksessa de Beer O.R. (toim.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, London: 287-303.
23) Strathmann R.R. (1975) Limits on diversity of forms: Piikkinahkaisten ambulakraalijärjestelmien haarautuminen. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: Functions and molecular adaptations. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Piikkinahkaisten toiminnallinen bauplan. Teoksessa Candia Carnevali M.D., Bonasoro F. (toim.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) Genes involved in the evolution of echinoderm morphology. Teoksessa Candia Carnevali M.D., Bonasoro F. (toim.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 232.
Share the post ”Veden suonet: The Evolution of the Echinoderm Water-Vascular System”
- Shares
 Kevin Zelnio (870 Posts)
Kevin Zelnio (870 Posts)