
Graphic used with permission. Daniel D. Brown, LaughingMantis.com.
Echinoderms are one of the most highly derived groups of animals with many species as a significant components of several marine communities.棘皮動物は、最も高度に派生した動物群のひとつです。 彼らは3つの基本的な共通の特徴によって分類されます。 1)5回対称、2)3次元の石灰質要素からなる骨格、3)外部に開口部を持つ水管系が存在すること、です。 特にこの水管系(WVS)は、水力システムのユニークな応用であり、数百の小さな愛らしい管足が凹凸のある表面を這い、ムール貝などの堅い殻の餌を食べるための圧力勾配を発生させることができる。
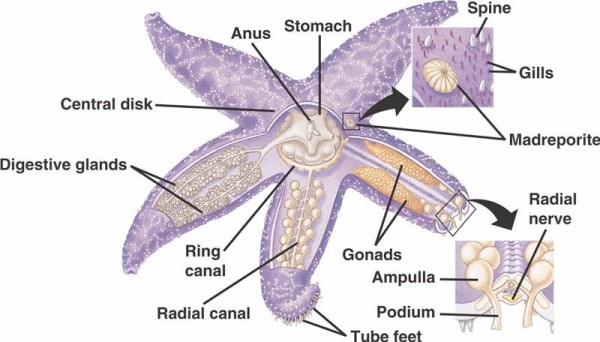
クリノイド以外の棘皮動物におけるWVS(以下略)の基本構成は、口腔周囲リング管とそこから伸びる5本の放射状の管から成っている。 また、環状運河には、海の外部環境と棘皮動物の内部環境との間のドアとして機能する心室突起が付着している。 マドレポライトは、マドレポラリアサンゴに似ていることから名づけられ、尾状の鞭毛をさまざまな方向に打ち出す細胞が並んでいる。WVS内部の液体の成分は海水に似ているが、カリウムイオンが最大60%多く、塩素イオンがやや高濃度で含まれ、自由に浮遊するアルバム状のタンパク質もある。 また、ナマコはWVS内に赤血球を持つことが確認されている。
各橈骨管は、管足に付着する短い管の対を連続的に繰り返しており、管足はその直上にある膨大部の筋収縮により作動する。 この管足は、管球の上にある筋肉が収縮することによって作動します。 また、管足は、橈骨(とうこつ)管に似た管から直接出ている3つのグループで発生します。

Xyloplax Voight 2005より。
棘皮動物の家系は完全には解明されていないが、カンブリア紀とオルドビス紀に短期間の多様化を経験し、その後基本的なバウプランはほとんど変化していない。 ウミユリ以外の4つのクラスは、そのWVSに類似した構造と機能を持つ。 ホロイトウ類は、バウプランの摂食触手が口腔周囲環状管から伸びており、骨格要素を含まないことから、クラスレベルでは最も派生的であると考えられる。 最近発見されたキシロプラックス(左)は、よりユニークなWVS配置をしており、径間接続を含む二重リングカナルと一続きの管足からなる新しいシステムである。 Janies and McEdward (1994)は、このユニークなWVSの形状は、幼生ベラトニクス小惑星の発生機構の変化により進化したことを示唆している。 このケースはWVSの最も現代的で派生した進化を表している。
棘皮動物は多様化が早く、保存状態が良くなく、不明瞭な化石群を比較するための複数のタイプ標本がないため、進化史を説明しようとすると問題が生じる。 WVSは厳密には棘皮動物だけの特徴ではない。 半索動物や水力で動く好塩基球の構造とも類似している。 また、棘皮動物の発生学的特徴は、脊索動物、半索動物、鞭毛虫の間でも共有されていることは言うまでもない。 さらに、環形孔はU字型の前駆体から発達し、やがて個体発生の過程で融合して、現在の棘皮動物の環を形成する。 これは、棘皮動物は幼生・成体ともにU字型の腸を持つlophophorateに似た先祖を共有していたというNichols (1967, 1972) の説と合致するように思える。
WVSは現在の棘皮動物ではユニークで、成功したイノベーションである。 この複雑な運河のネットワークは、ophophateのような共通祖先のU字型腸のような、既存の解剖学的構造の上に構築された可能性がもっともらしいが、その証拠は不明である。 私にとって印象的なのは、このようなイノベーションが他の動物門に存在しないことである。 WVSはおそらく、化石の解釈から明らかなように、初期の無柄棘皮動物で摂食構造として生じ、二次的に運動にも使われるようになったのであろう。 また、絶滅した棘皮動物や現存する棘皮動物の間で、WVSが異なる目的(呼吸、摂食、運動)のために複数回進化したという解釈もできるかもしれない。 いずれにせよ、WVSは棘皮動物の中でも驚くべき適応である。

From Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) New Zealand A new class of Echinodermata from New Zealand. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) 古生代小惑星の水管系と機能形態. Lethaia 21: 189-206.
3)ビニヨンJ.(1972)棘皮動物の生理. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) ヒトデの水管系作動に伴う体液損失の再評価. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrates. シナウアー・アソシエイツ, INC. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7)ハイマン L. (1955) 無脊椎動物: 第四巻 棘皮動物.
8) Janies D.A., McEdward L.R. (1993) Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development.棘皮動物における棘皮動物および水生血管の高次構造. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) 同心円状の水脈血管系の進化に関する仮説. ウィルソン・ジュニア W.H.・ストリッカー S.A.・シン G.L. (eds.) による。 海産無脊椎動物の生殖と発生. 10) Jeffries R.P.S. (1988) How to characterize the Echinodermata -Some implications of the sister-group relationship between echinoderms and chordates. ポール C.R.C., スミス A.B. (eds.) Echinodermy Phylogeny and Evolutionary Biology.による。 オックスフォード大学出版局,ロンドン:1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) 脊索動物と棘皮動物の初期系統と脊索動物の左右対称性と両側対称性の起源. Acta. Zool. 77: 101-122.
12) ローレンス J.M. (1987) A Functional Biology of Echinoderms. Croom Helm Ltd.,ロンドン.
13) Lowe C.J., Wray G.A. (1997) 棘皮動物の進化におけるホメオボックス遺伝子の役割の根本的な変化. ネイチャー389: 718-721.
14) メンゲ B.A. (1982) 摂食の環境に対する影響. 小惑星の生物. Jangoux M.・Lawrence J.M. (eds.) による。 棘皮動物栄養学. A.A. Balkema, Rotterdam: 521-552.
15)ニコルス D. (1967)棘皮動物の起源. ミロットN. (編). 棘皮動物の生物学. (1972)棘皮動物における水脈系. 古生物学15(4):519-538.
17) ポール C.R.C. (1977) 原始棘皮動物の進化. ハラムA.編. 化石記録にみる進化のパターン. 18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19)。 Phelan T.F. (1977) Clypeasteroid Echinoids の水管系、食物溝、祖先に関するコメント. Bull. Mar. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) ヒトデ水管系における体液量の維持. Nature 262: 577-578.
21) Smith A.B. (1988) To group or not to group: Xyloplaxの分類学的位置づけ. Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (eds.) による。 棘皮動物の生物学. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Some aspects of evolution in Echinodermata. de Beer O.R. (ed.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, London: 287-303.
23)ストラスマン R.R. (1975) 形態の多様性の制限: 棘皮動物の外肛門系の分岐. アメリカ. Nat. 24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: 機能および分子的適応. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) 棘皮動物の機能性バウブラン. キャンディア・カルネヴァリM.D.・ボナソロF.編: 棘皮動物研究1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) 棘皮動物の形態進化に関与する遺伝子. キャンディア・カーネヴァリM.D.・ボナソロF.編. 棘皮動物研究1998. A.A. Balkema, Rotterdam: 232.
Share the post “Veins of Water: The Evolution of the Echinoderm Water-Vascular System”
- Shares
 Kevin Zelnio (870 Posts)
Kevin Zelnio (870 Posts) の投稿を公開しました。