
Graphic used with permission. Daniel D. Brown, LaughingMantis.com.
Echinodermen zijn een van de meest afgeleide groepen dieren met vele soorten als belangrijke componenten van diverse mariene gemeenschappen. Ze worden geclassificeerd door drie fundamentele gemeenschappelijke kenmerken: 1) pentaradiale symmetrie, 2) skelet bestaande uit driedimensionale calcietelementen, en 3) de aanwezigheid van een watervatenstelsel met een uitwendige opening. Met name dit watervatenstelsel (WVS) blijft een unieke toepassing van een hydraulisch systeem, dat drukgradiënten genereert die in staat zijn honderden piepkleine, adorabele buisvoetjes te bevelen om over oneffen oppervlakken te kruipen en zich te voeden met taaie prooien met schelpen, zoals mosselen.
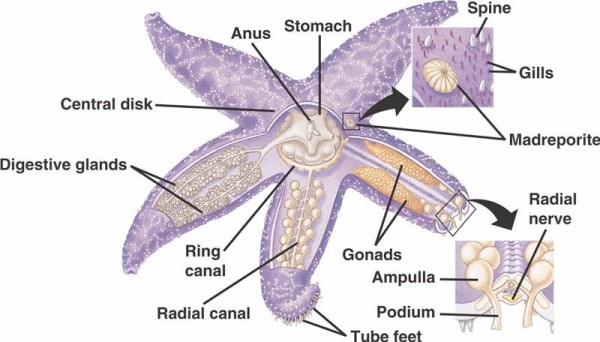
De basiscomponenten van het WVS bij niet-crinoide stekelhuidigen (hieronder) omvatten een circumoraal ringkanaal met vijf radiaalkanalen die zich van daaruit uitstrekken. Aan het ringkanaal is ook de madreporiet bevestigd, die fungeert als een deur tussen het externe milieu van de zee en het interne milieu van het stekelhuidje. De madreporiet, genoemd naar zijn gelijkenis met het Madreporaria koraal (wat “moeder van de poriën” betekent), is bekleed met cellen die staartachtige flagellen in verschillende richtingen slaan.De vloeistof binnenin de WVS is qua samenstelling vergelijkbaar met zeewater, maar bevat tot 60% meer kalium-ionen, iets hogere concentraties chloride-ionen, en vrij zwevende albumine-achtige proteïnen. Bij zeekomkommers zijn ook rode bloedcellen in hun WVS aangetroffen.
Elk radiaal kanaal heeft zich in serie herhalende paren kortere kanalen die vastzitten aan de buisvoeten, die worden bediend door spiersamentrekking van de ampullen die zich direct boven hen bevinden. Denk aan een kalkoenbeker: als je in de bol knijpt, duwt hij vloeistof uit de bol, als je de druk loslaat, zuigt hij vloeistof terug naar binnen, naar de bol toe. Crinoïden verschillen van elkaar omdat zowel deze kortere kanalen als de ampullae ontbreken en hun buisvoeten in groepjes van drie voorkomen die rechtstreeks uit een kanaal komen dat lijkt op het radiale kanaal.
Crinoïden behouden de meest primitieve vorm van de WVS. Men dacht dat de buisvoeten oorspronkelijk volledig werden gebruikt voor de ademhaling en als voedingsstructuren, maar fossiele stekelhuidigen hebben duidelijke poriën tussen de skeletplaten, waaruit Nichols (1972) concludeert dat het buisvoetsysteem niet zeer uitgebreid was in het begin van de crinoide evolutie. Maar hij laat andere interpretaties van de structuren buiten beschouwing, zoals openingen voor inwendige onderdelen die te maken hebben met voortbeweging of voeding en die geen verband houden met ademhaling. Carpoïden, zoals Gyrocystis en Dendrocystites, die in afzettingen uit het midden van het Cambrium worden gevonden, worden beschouwd als basaal voor de lijn van de stekelhuidigen, vanwege de gelijkenis in skeletelementen, en er wordt verondersteld dat zij zachte, uittrekbare inwendige tentakelstructuren hebben. Het bestaan van een primitief WVS is niet onomstotelijk bewezen in de fossiele gegevens van de carpoïden, maar toch wordt aangenomen dat het zich vóór andere kenmerken van de stekelhuidigen heeft ontwikkeld. De vroegste suggestie van een WVS is afkomstig van de Helicoplacoidea, gevonden in afzettingen uit het lagere Cambrium (gekenmerkt door de aanwezigheid van sterk georganiseerde rijen ambulacrale poriën). Volgens Paul (1977) is er geen moderne klasse die is afgeweken van de carpoïden, die sinds het Mesozoïcum zijn uitgestorven. Helicoplacoïden, met triradiale symmetrie, divergeerden zeer vroeg van de carpoïden en de rest van de echinodermische lijn evolueerde op het Helicoplacoïd bauplan.

Xyloplax van Voight 2005.
Hoewel de stamboom van de stekelhuidigen niet volledig is uitgewerkt, hebben zij een korte diversificatie doorgemaakt tijdens het Cambrium en het Ordovicium en is het basisbauplan sindsdien weinig veranderd. De vier niet-crinodermenklassen hebben een soortgelijke structuur en functie in hun WVS. De Holothuroiden zijn waarschijnlijk het meest afgeleid op klassenniveau, omdat hun bucchale voedingstentakels vanuit het circumorale ringkanaal zijn verlengd en geen skeletelementen bevatten. De recent ontdekte Xyloplax (links), een vreemd soort zeester, heeft een meer unieke WVS opstelling: een nieuw systeem met dubbele ringkanalen met inter-radiale verbindingen en mono-seriële set van buisvoetjes. Janies en McEdward (1994) suggereren dat de unieke geometrie van zijn WVS is geëvolueerd via een wijziging in het ontwikkelingsmechanisme van een juveniele velatide zeester. Dit geval vertegenwoordigt de meest moderne en afgeleide evolutie van de WVS.
De snelle diversificatie van de stekelhuidigen, het gebrek aan goede conservering en het ontbreken van meer dan één type specimen voor het vergelijken van obscure fossiele klassen levert problemen op bij het trachten te verklaren van hun evolutionaire geschiedenis. De WVS is, strikt genomen, niet alleen een uniek kenmerk van stekelhuidigen. Er zijn overeenkomsten met structuren in de hemichordaten en de hydraulisch bediende lophophoor. Niet te vergeten dat embryologische kenmerken van stekelhuidigen ook gedeeld worden door chordaten, hemichordaten, en de lophophoraten. Bovendien heeft het ringkanaal zich ontwikkeld uit een U-vormige voorloper die uiteindelijk tijdens de ontogenie samensmelt om de ring te vormen die we nu bij stekelhuidigen zien. Dit lijkt te passen in Nichols’ (1967, 1972) theorie dat stekelhuidigen een voorouder hebben gedeeld met een lophophoraatachtige voorganger, die ook U-vormige ingewanden hebben in zowel larvale als volwassen vormen.
Het WVS is een unieke en, in de huidige stekelhuidigen, succesvolle innovatie. Het lijkt aannemelijk dat het ingewikkelde netwerk van kanalen kan zijn voortgebouwd op reeds bestaande anatomische structuren, zoals de lophophoor en de U-vormige darm van een lophophate-achtige gemeenschappelijke voorouder, maar het bewijs daarvoor is onduidelijk. Wat mij opvalt, is dat een dergelijke innovatie niet voorkomt in andere fyla. De WVS is waarschijnlijk ontstaan als een voedingsstructuur in vroege sessiele stekelhuidigen, zoals blijkt uit fossiele interpretaties, en werd in tweede instantie gebruikt voor voortbeweging. Een andere interpretatie kan zijn dat de WVS meer dan eens geëvolueerd is onder uitgestorven en levende stekelhuidigen voor verschillende doeleinden (d.w.z. ademhaling, voeding, voortbeweging). Hoe dan ook, de WVS is een verbazingwekkende aanpassing onder de Echinodermata.

Van Paul & Smith 1984
1) Baker A.N., Rowe F.W.E., Clark H.E.S. (1986) A new class of Echinodermata from New Zealand. Nature 321: 862-864.
2) Blake D.B., Guensburg T.E. (1988) The water vascular system and functional morphology of Paleozoic asteroids. Lethaia 21: 189-206.
3) Binyon J. (1972) Physiology of Echinoderms. Pergamon Press Ltd., Oxford.
4) Binyon J. (1984) A reappraisal of the fluid loss resulting from operation of the water vascular system of the starfish, Asterias rubens. J. Mar. Biol. Ass. UK 64(3): 726.
5) Brusca R.C., Brusca G.J. (2003) Invertebrates. Sinauer Associates, Inc. Sunderland, MA.
6) Durham J.W. (1964) The Helioplacoidea and some possible implications. Yale Scient. Mag. 39(2): 24-28.
7) Hyman L. (1955) The Invertebrates: Vol. IV Echinodermata. McGraw-Hill, New York.
8) Janies D.A., McEdward L.R. (1993) Highly derived coelomic and water-vascular morphogensis in a starfish with pelagic direct development. Biol. Bull. 185: 56-76.
9) Janies D.A., McEdward L.R. (1994) A hypothesis for the evolution of the Concentricycloid water-vascular system. In Wilson Jr. W.H., Stricker S.A., Shinn G.L. (eds.): Reproduction and Development of Marine Invertebrates. John Hopkins University Press, Baltimore.
10) Jeffries R.P.S. (1988) How to characterize the Echinodermata-s some implications of the sister-group relationship between echinoderms and chordates. In Paul C.R.C., Smith A.B. (eds.) Echinoderm Phylogeny and Evolutionary Biology. Oxford University Press, Londen: 1-13.
11) Jeffries R.P.S., Brown N.A., Daley P.E.J. (1996) The early phylogeny of chordates and echinoderms and the origin of chordate left-right symmetry and bilateral symmetry. Acta. Zool. 77: 101-122.
12) Lawrence J.M. (1987) A Functional Biology of Echinoderms. Croom Helm Ltd., Londen.
13) Lowe C.J., Wray G.A. (1997) Radical alterations in the roles of homeobox genes during echinoderm evolution. Nature 389: 718-721.
14) Menge B.A. (1982) Effecten van voeding op de omgeving: Asteroidea. In Jangoux M., Lawrence J.M. (eds.): Echinoderm Nutrition. A.A. Balkema, Rotterdam: 521-552.
15) Nichols D. (1967) The origin of echinoderms. In Millott N. (ed.): Echinoderm Biologie. Academic Press Inc., London: 240-255.
16) Nichols D. (1972) The water-vascular system in living and fossil echinoderms. Paleontologie 15(4): 519-538.
17) Paul C.R.C. (1977) Evolution of primitive echinoderms. In Hallam A. (ed.): Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam: 123-158.
18) Paul C.R.C., Smith A.B. (1984) The early radiation and phylogeny of echinoderms. Biol. Rev. 59: 443-481.
19) Phelan T.F. (1977) Comments on the water vascular system, food grooves, and ancestry of Clypeasteroid Echinoids. Bull. Mar. Sci. 27(3): 400-422.
20) Prusch R.D., Whoriskey F. (1976) Handhaving van het vloeistofvolume in het watervatenstelsel van zeesterren. Nature 262: 577-578.
21) Smith A.B. (1988) To group or not to group: De taxonomische positie van Xyloplax. In Burke R.D., Mladenov P.V., Lambert P., Parsley R.L. (eds.): Echinoderm Biology. A.A. Balkema, Rotterdam,17-23.
22) Spencer W.K. (1938) Some aspects of evolution in Echinodermata. In de Beer O.R. (ed.): Evolution, Essays and Aspects of Evolutionary Biology Presented to E.S. Goodrich. Oxford University Press, London: 287-303.
23) Strathmann R.R. (1975) Beperkingen van de vormenrijkdom: Branching of ambulacral systems of echinoderms. Amer. Nat. 109: 177-190.
24) Turbeville J.M., Schulz J.R., Raff R.A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evo. 11: 648-655.
25) Voight, J.R. (2005) First Report of the Enigmatic Echinoderm Xyloplax from the North Pacific. Biol. Bull. 208(2): 77-80.
26) Weber R.E., Vinogradov S.N. (2001) Nonvertebrate hemoglobins: Functies en moleculaire aanpassingen. Phys. Rev. 81(2): 569-628.
27) Welsch U. (1998) Functional bauplan of the echinoderms. In Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 39-47.
28) Wray G.A. (1999) Genes involved in the evolution of echinoderm morphology. In Candia Carnevali M.D., Bonasoro F. (eds.): Echinoderm Research 1998. A.A. Balkema, Rotterdam: 232.
Deel het bericht “Aderen van water: The Evolution of the Echinoderm Water-Vascular System”
- Shares
 Kevin Zelnio (870 Posts)
Kevin Zelnio (870 Posts)